








方案详情
文
采用LaVison公司的粒子成像测速系统(PIV),测量了模拟水池中模拟肩并肩游泳运动员同步运动模型的流场。
方案详情

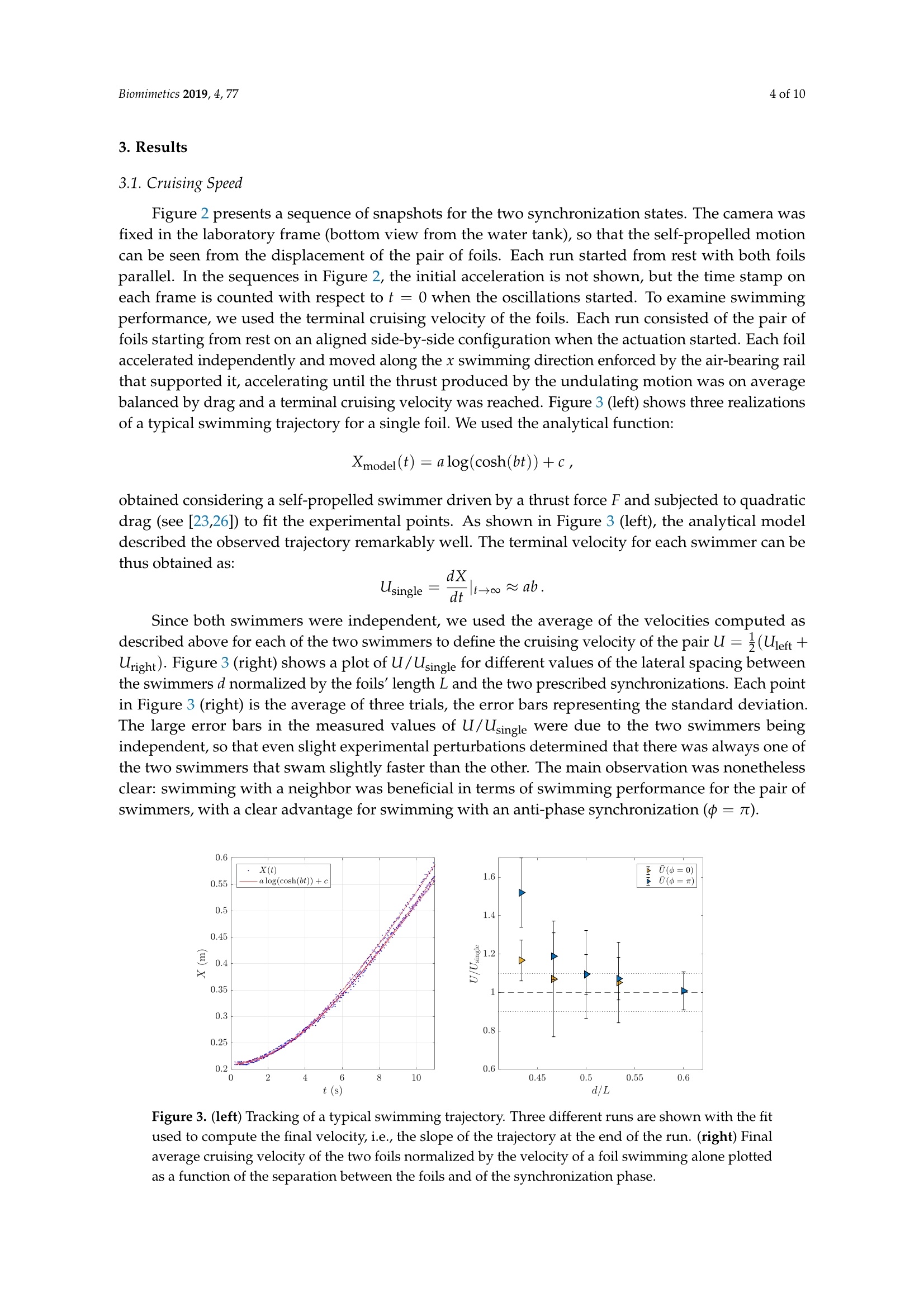
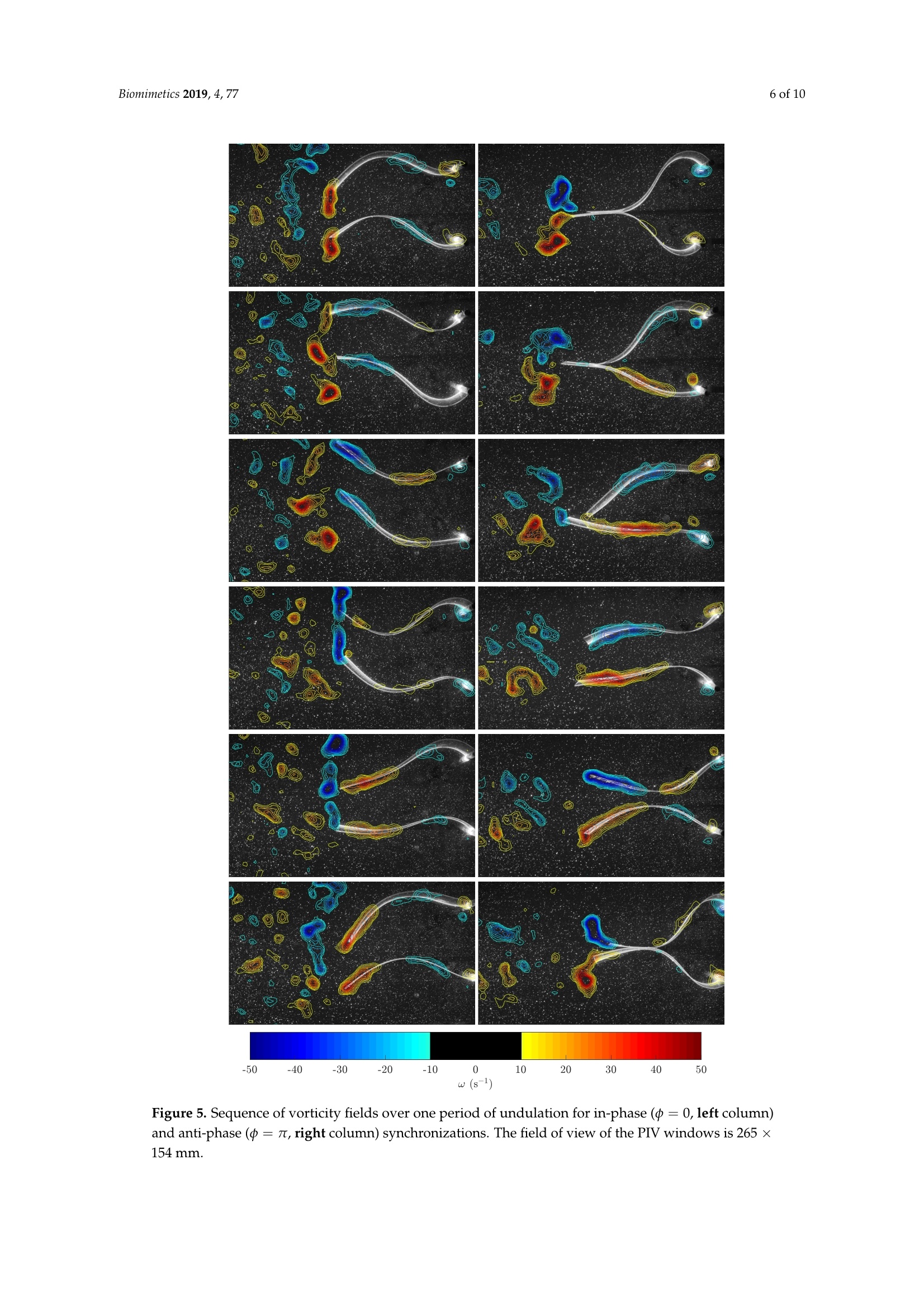
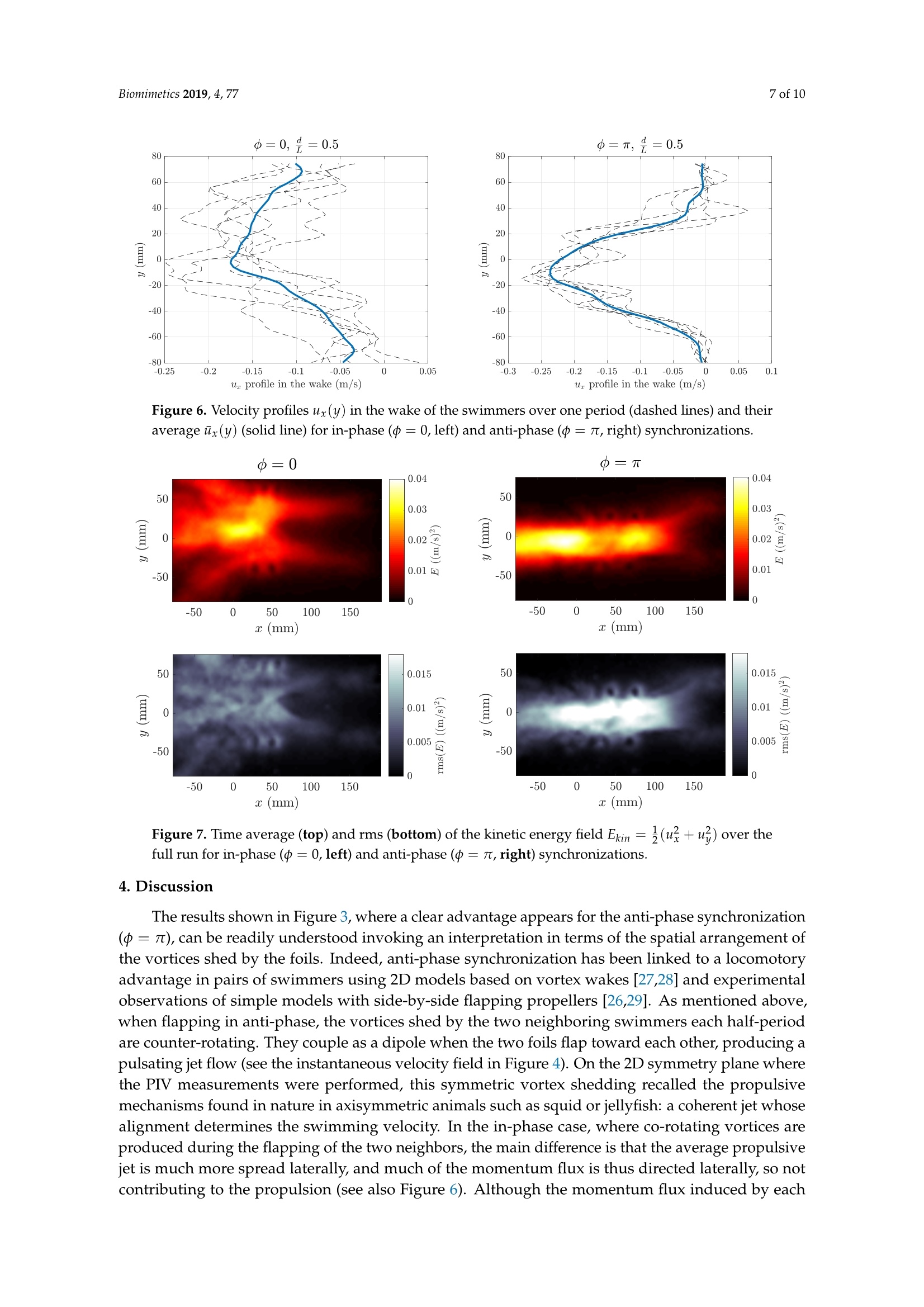
biomimetics Biomimetics 2019, 4, 772 of 10 Biomimetics 2019,4,77; doi:10.3390/biomimetics4040077www.mdpi.com/journal/biomimetics Article On the Fluid Dynamical Effects of Synchronization inSide-by-Side Swimmers Ramiro Godoy-Diana**iD, Jerome Vacher, Veronica Raspa fippaand Benjamin ThiriaD Laboratoire de Physique et Mecanique des Milieux Héterogenes (PMMH, UMR 7636), CNRS, ESPCI Paris-PSLUniversity, Sorbonne Universite, Universite de Paris, F-75005 Paris, France; jerome.vacher@espci.org (J.V.);veronica.raspa@gmail.com (V.R.); bthiria@pmmh.espci.fr (B.T.) 米 Correspondence: ramiro@pmmh.espci.fr 十 Current address: Departamento de Fisica, Facultad de Ciencias Exactas y Naturales, Universidad de BuenosAires, IFIBA-CONICET,Ciudad Universitaria, Buenos Aires 1428, Argentina. Received: 28 August 2019; Accepted:2 December 2019; Published:5 December 2019 check forupdates Abstract: In-phase and anti-phase synchronization of neighboring swimmers is examined experimentallyusing two self-propelled independent flexible foils swimming side-by-side in a water tank. The foilsare actuated by pitching oscillations at one extremity-the head of the swimmers-and the flowengendered by their undulations is analyzed using two-dimensional particle image velocimetry intheir frontal symmetry plane. Following recent observations on the behavior of real fish, we focus onthe comparison between in-phase and anti-phase actuation by fixing all other geometric and kinematicparameters. We show that swimming with a neighbor is beneficial for both synchronizations tested,as compared to swimming alone, with an advantage for the anti-phase synchronization. We showthat the advantage of anti-phase synchronization in terms of swimming performance for the two-foil“school" results from the emergence of a periodic coherent jet between the two swimmers. Keywords: collective swimming; bio-inspired swimmers; fluid-structure interaction; self-propulsion;synchronization; fish schooling 1. Introduction The interaction between neighboring individuals constitutes the fundamental fabric of collectivedynamics [1]. Depending on the system, this interaction can be established by many different kinds ofsensing and actuation, involving for instance optical, chemical, magnetic, or mechanical signals [2-4],and the collective response can emerge from nonlinear feedbacks based on local interactions [5]. In thecase of swimmers (see, e.g., [6] for a review on sensing and actuation in fish inspired robots), biologicalor artificial, regardless of the kind of sensing, their interaction is mediated by hydrodynamics.The understanding of interactions between neighbors is not only crucial for the understanding of fishschooling,but also in the case of robotic swimmers, where one would like to have accurate modelsor design guidelines for the optimal behavior of a robot in the presence of neighbors. Concerningfish schools, it is challenging to include hydrodynamic interactions in models with large numbers ofswimmers without resorting to simplified theoretical approximations, e.g.,[7,8]. On the other hand,zooming into the problem of a single swimmer, despite the successes of Lighthill's theory to describethe mechanics of fish swimming [9], the ubiquitous problems of the interaction between a swimmerand its environment are out of its reach. This environment can be a wall, a substrate (a significantnumber of works have explored this problem in different contexts [10-13]), or another neighboringswimmer. The interactions between multiple swimmers may significantly impact the performanceor cost of locomotion associated with fish schooling, as each swimmer moves in a non-uniform andunsteady flow created by its neighbors. Collective effects in fish swimming have been widely studied theoretically/numerically, e.g., [14-16], and experimentally [17-19]. The goal of the present workis to examine the hydrodynamics of a basic experiment of the interaction between two neighboringswimmers, which we model as undulating flexible foils. Actuated flexible foils have been extensively studied in the last decade as a model of fish-likeswimmers or as a bio-inspired propeller, and their dynamical behavior is ruled by a complex fully-coupled fluid-structure interaction problem, e.g., [20-22]. In this paper, we report on an experimentwith two self-propelled independent swimmers, each one constituted by a flexible foil actuated bypitching oscillations at one extremity, placed in a side-by-side configuration with imposed swimmingdirection (see Figure 1). We have already used different versions of the experiment described here,on the one hand to examine the dynamics of a single swimmer [22,23], and on the other hand to considerthe effect of swimming near a wall [12]. In the latter, we established the main physical mechanism thatleads to a performance enhancement due to the presence of the wall: a reorientation of momentum. Conceptually, the setup with two swimmers that we study in the present paper is one of the simplestmodel realizations of the minimal school [24,25] and has been designed to examine the complex flowdynamics that arises due to the undulation kinematics of the two swimmers. Recent experiments on realfish in a swimming channel with imposed velocity have shown that fish favor a synchronized kinematicswith their nearest neighbors as the swimming speed of the school increases [17,18]. That observation hasbrought evidence to previous numerical predictions on the hydrodynamic advantages of synchronizedswimming [16] and is the seed for the specific objective of the present paper: to compare the flowdynamics and the performance of the pair of interacting swimming foils in the two possible states ofsynchronization: in-phase and anti-phase kinematics. In order to focus on this comparison, we pickeda single point in the kinematic, geometric, and material parameter space for the case of an elastic foilactuated by pitching oscillations, thus fixing several parameters that define the problem. Figure 1. Schematic diagrams of the experiment with two self-propelled swimmers in a side-by-sideconfiguration. Each foil is held by an independent air-bearing rail (left) and actuated by an oscillatingpitching motion of an axis that holds the foil from one extremity (right). 2. Experimental Setup Following [23], the experiments were carried out in a free-surface water tank (0.9 m x0.8 m ×0.5 m) with no external flow where two swimmers were fully submerged and allowed to move freelyin a rectilinear motion, independently, using an air-bearing rail to hold each swimmer (see Figure 1).Each swimmer was made of a thin rectangular foil (length L =0.15 m, span W=0.1 m, and thickness130 um, made of Mylar) of bending rigidity B=1.02×10-3Nm. The foil was clamped to a cylindricalaxis (d=0.005 m in diameter) that constituted the head of the swimmer and set the swimming depthin the water bulk. A pitching motion was imposed, which generated self-propulsion by creating abackward propagating undulation along the flexible body. The rotational oscillation of the head of each swimmer was controlled with 0.5 degrees of precision by a small stepper (see Figure 1) and a sinewave driving curve so that: The amplitude of the oscillation was fixed for the present experiments at 0o =60°, while thefrequency was set at f = 2 Hz (except for the visualization of Figure 2, where the frequency wasf=3Hz). The main parameter examined was the synchronization state, determined by the value of中 in the actuation signal: 中 =0 corresponds to the two neighboring swimmers undulating in-phase,whereas 中= n corresponds to the anti-phase case. We also varied the side-by-side distance d betweenthe two neighboring swimmers over a small range d/L E [0.4,0.6]. With the aforementioned actuation,the Reynolds number Re=UL/v based on the foil length L and the self-propelled terminal swimmingspeed U (v being the kinematic viscosity of the fluid) was Re ≈ 7500, and the Strouhal number basedon the tip-to-tip amplitude at the tail StA=fA/U~ 1. Figure 2. Two image sequences are presented with the two foils actuated in-phase (top) and in anti-phase (bottom). The camera is placed at a fixed position below the water tank, and the swimmingdirection is directed toward the top of the figure. Time in these frame sequences goes from left to right,with an interval between frames of 0.24 s. The initial acceleration phase is not shown. The time stamp oneach frame is counted with respect tot=0when the oscillations start from rest. The case shown herecorresponds to d/L=0.6 and f=3Hz. The size of the visualization window is approximately L×2L(0.15×0.3m). Particle Image Velocimetry PIV measurements were performed using a LaVision system (DaVis 7.2) with a QuantronixDarwin-Duo Nd:YLF double-cavity pulsed laser (20 mJ, 527 nm) and a Phantom V9 camera at fullresolution: 1632 ×1200. Image acquisition was performed at 100 Hz during 10 s, which was enoughtime for the self-propelled swimmers to accelerate and leave the measurement window (265×154 mm).The laser sheet (of thickness ≈1 mm) was placed at mid-span of the foils and illuminating the foilfrom the trailing edge. The PIV computation was performed using a multi-pass algorithm with finalinterrogation windows of 32 ×32 pixels? and 50% overlap. A standard median filter to remove badvectors and replace them with second or third correlation peaks was used, and holes in the vectorfield were filled by interpolation before computing the vorticity fields. Data analysis, post-processing,and visualization were performed using MATLAB and the PIVMat toolbox. 3. Results 3.1. Cruising Speed Figure 2 presents a sequence of snapshots for the two synchronization states. The camera wasfixed in the laboratory frame (bottom view from the water tank), so that the self-propelled motioncan be seen from the displacement of the pair of foils. Each run started from rest with both foilsparallel. In the sequences in Figure 2, the initial acceleration is not shown, but the time stamp oneach frame is counted with respect to t = 0 when the oscillations started. To examine swimmingperformance, we used the terminal cruising velocity of the foils. Each run consisted of the pair offoils starting from rest on an aligned side-by-side configuration when the actuation started. Each foilaccelerated independently and moved along the x swimming direction enforced by the air-bearing railthat supported it, accelerating until the thrust produced by the undulating motion was on averagebalanced by drag and a terminal cruising velocity was reached. Figure 3 (left) shows three realizationsof a typical swimming trajectory for a single foil. We used the analytical function: obtained considering a self-propelled swimmer driven by a thrust force F and subjected to quadraticdrag (see [23,26]) to fit the experimental points. As shown in Figure 3 (left), the analytical modeldescribed the observed trajectory remarkably well. The terminal velocity for each swimmer can bethus obtained as: Since both swimmers were independent, we used the average of the velocities computed asdescribed above for each of the two swimmers to define the cruising velocity of the pair U=(Uieft+Uright). Figure 3 (right) shows a plot of U/Usingle for different values of the lateral spacing betweenthe swimmers d normalized by the foils’ length L and the two prescribed synchronizations. Each pointin Figure 3 (right) is the average of three trials, the error bars representing the standard deviation.The large error bars in the measured values of U/Usingle were due to the two swimmers beingindependent, so that even slight experimental perturbations determined that there was always one ofthe two swimmers that swam slightly faster than the other. The main observation was nonethelessclear: swimming with a neighbor was beneficial in terms of swimming performance for the pair ofswimmers, with a clear advantage for swimming with an anti-phase synchronization (中= 元). Figure 3. (left) Tracking of a typical swimming trajectory. Three different runs are shown with the fitused to compute the final velocity, i.e., the slope of the trajectory at the end of the run. (right) Finalaverage cruising velocity of the two foils normalized by the velocity of a foil swimming alone plottedas a function of the separation between the foils and of the synchronization phase. 3.2. Flow Field Measurements We examine now the flow field in the horizontal (xy) symmetry plane of the swimmers at mid-span(z= W/2), using the output of the PIV measurements u(x,y)=(ux,Uy). A snapshot of the velocityfield for a single swimmer and the equivalent cases for a pair of swimmers in in-phase (中=0) andanti-phase (中=nt) synchronizations is presented in Figure 4, together with the corresponding fieldsof vorticity w=Vxu. In the single swimmer case, the velocity field showed the backward jet flowassociated with thrust production, while the vorticity field showed the cases with two swimmers. It isremarkable that the two different synchronizations produced very different flow fields: the in-phasecase can be qualitatively understood as the superposition of two single swimmers, which could beexpected; at the time chosen for the snapshot in Figure 4, each foil was shedding a vortex rotatingcounterclockwise, and the backward jet was just more spread laterally than the case of the singleswimmer. On the contrary, in the anti-phase case, the interaction between the flow produced by thetwo foils intensified dramatically the backward jet (see the velocity field), and the vortex sheddingwas symmetrized by the simultaneous shedding of a couple of counter-rotating vortices (see thevorticity field). 3 Figure 4. (Top row) Snapshot of a typical instantaneous velocity field for a single swimmer (left) anda pair of swimmers in phase (中=0) and anti-phase (中=n) (middle and right frames, respectively).(Bottom row) Corresponding vorticity fields. The field of view of the PIV windows is 265 × 154 mm. In order to see the time evolution of the flow structures, Figure 5 presents a sequence of snapshotsof the vorticity field over one period of undulation for in-phase(中=0) and anti-phase (中=7t)synchronizations (the corresponding videos are available as Supplementary Material). As mentionedabove, the main vortices shed at the trailing edge with each oscillation were co-rotating in the in-phasecase and counter-rotating in the anti-phase case. The main effect in the in-phase case was an undulatingflow field that widened the lateral extension of the wake, whereas in the anti-phase case, the twocounter-rotating vortices produced by the two neighboring swimmers gave rise to a coherent pulsatingiet flow between them. A global quantitative picture of the previous flow features is presented in Figures 6 and 7, which show,respectively, a sequence of ux(y) profiles measured just behind the swimmer together with theiraverage profile ux(y) over one period and a time average of the kinetic energy Ekin=(u?+u) forthe full run, for the in-phase and anti-phase cases of Figure 5. Figure 5. Sequence of vorticity fields over one period of undulation for in-phase (中=0, left column)and anti-phase(中=, right column) synchronizations. The field of view of the PIV windows is 265 ×154 mm. u profile in the wake (m/s) Figure 6. Velocity profiles ux(y) in the wake of the swimmers over one period (dashed lines) and theiraverage ux(y) (solid line) for in-phase (中=0, left) and anti-phase(中= 7t, right) synchronizations. o=0 中=T Figure 7. Time average (top) and rms (bottom) of the kinetic energy field Ekin =2(u+uh) over thefull run for in-phase (中=0,left) and anti-phase (中=t, right) synchronizations. 4. Discussion The results shown in Figure 3, where a clear advantage appears for the anti-phase synchronization(中= n), can be readily understood invoking an interpretation in terms of the spatial arrangement ofthe vortices shed by the foils. Indeed, anti-phase synchronization has been linked to a locomotoryadvantage in pairs of swimmers using 2D models based on vortex wakes [27,28] and experimentalobservations of simple models with side-by-side flapping propellers [26,29]. As mentioned above,when flapping in anti-phase, the vortices shed by the two neighboring swimmers each half-periodare counter-rotating. They couple as a dipole when the two foils flap toward each other, producing apulsating jet flow (see the instantaneous velocity field in Figure 4). On the 2D symmetry plane wherethe PIV measurements were performed, this symmetric vortex shedding recalled the propulsivemechanisms found in nature in axisymmetric animals such as squid or jellyfish: a coherent jet whosealignment determines the swimming velocity. In the in-phase case, where co-rotating vortices areproduced during the flapping of the two neighbors, the main difference is that the average propulsivejet is much more spread laterally, and much of the momentum flux is thus directed laterally, so notcontributing to the propulsion (see also Figure 6). Although the momentum flux induced by each independent swimmer was on average the same, the phase difference ruled the interference betweenthe propulsive vortex streets produced by each swimmer, determining that flapping in anti-phase wasmore effective for the pair. A global view of the different effects obtained by in-phase and anti-phasesynchronization can be seen by examining the kinetic energy in the area traversed by the swimmers(see Figure 7). The "hot"zones in this average picture are clearly different in the two cases: a spreadwake in the in-phase case vs. a coherent energetic zone in the anti-phase case. In spite of the figuresshowing an average field, the unsteadiness of the flow field can be seen in the outer parts of the wake,where the signature of each foil oscillation and its associated vortex shedding is visible, especially inthe rms fields. The mechanisms described above have been invoked as a possible explanation for the slightpreference for anti-phase synchronization observed in pairs of tetra fish as their swimming velocityincreases [17], but without any velocimetry measurement in the flow field between the fish. In additionto synchronization, the other crucial parameter was the position of the two neighboring swimmerswith respect to each other, which would determine the spatial pattern of the fish school in larger groups.The aforementioned experiments were performed on a shallow swimming channel such that the groupof fish adopted a quasi-two-dimensional spatial arrangement. With the same setup, Ashraf et al. [18]showed that the synchronized swimming, in-phase and anti-phase, observed when fish are swimmingfast is accompanied by a spatial arrangement where nearest neighbors adopt a phalanx pattern andare thus closer to each other, with a peak in the nearest-neighbor distance (NND) lower than 0.5 bodylengths. The effect of the spatial configuration on swimming performance has also been investigatedexperimentally in a model system considering the propulsive performance of a pair of pitching foils instaggered configurations [29]. The preference of fish for such patterns where nearest neighbors are close to each other and theirkinematics are synchronized supports a two-dimensional physical interpretation: in the in-phase casebased on the channeling effect [16,27] and in the anti-phase case based on the pulsating jet mechanismdescribed here with the two foil experiment. Indeed, 2D models have been successful in describingother schooling configurations; see, e.g.,[30]. However, the thrust production, energy expenditure,and stability of a fish school are very sensitive to the details of the pattern, as shown recently byLi et al. [24] using 3D numerical simulations. Although 2D models and interpretations such as the onewe have used to explain the advantage of the anti-phase synchronization can be useful to understandthe basic physical mechanisms at play during schooling, the 3D reality needs to be always kept inmind, especially in a bio-inspired robotics perspective. Author Contributions: Conceptualization, R.G.-D. and B.T.; methodology, R.G.-D., J.V., V.R., and B.T.; software,J.V. and V.R.; validation, R.G.-D. and J.V.; investigation, R.G.-D., J.V., V.R., and B.T.; writing, original draftpreparation, R.G.-D.; writing, review and editing, R.G.-D., V.R., and B.T.; supervision, R.G.-D. and B.T. Funding: This research received no external funding. Acknowledgments: We acknowledge support of the French-Argentinian International Research Project IVMF,CNRS-INSIS (France), CONICET (Argentina). Conflicts of Interest: The authors declare no conflict of interest. Abbreviations The following abbreviations are used in this manuscript: PIV particle image velocimetry NND nearest-neighbor distance 2D two-dimensional 3D three-dimensional ( References ) ( 1. Vicsek,T.; Zafeiris, A. Collective motion. P h ys. Rep. 2012,517,71-140. [Cro s s R e f] ) ( 2. Kube, C.R.; Zhang, H. C o llective robotics: From s o cial insects to robots. Adapt. Behav. 1993,2,189-218. C rossRe f ) ( 3. Popat, R .; Cornforth, D .M.; M c Nally, L. ; Br o wn, S.P . Co l lective sen s ing and coll e ctive responses inquorum-sensing bacteria. J. R. Soc . Interface 2015,12,20140882. [CrossR e f] [ PubMed] ) ( 4. Ramdya,P.; Lichocki, P.; Cruchet, S.; Frisch, L.; Tse, W . ;Floreano , D.; Benton, R. Mechanosensory interactionsdrive collective behavior in Drosophila. Nature 2015,519,2 3 3. [C rossRef] [P ub M ed] ) ( 5. Halloy, J.; Sempo, G . ; Caprari, G.; Rivault, C. ; Asadpour, M.; T a che, F.; Said, I.; Durier, V.; C a nonge, S.; Ame, J.M.; et al. Social integration of robots i n to groups of cockroaches to control self-organized choices. Science 2007, 318, 1155-1158. [CrossRef ][ PubMed] ) ( 6. Raj, A.; Thakur, A. Fish-inspire d robots: Design,sensing, actuation, and autonomy-A review of research. Bioinspir. Biomim. 2016,11,1-23. [ C rossR e f] ) ( 7. Lopez, U.; Gautrais,J.; Couzin, I.D. ; Theraulaz,G. From behavioral analyses to models of collective motionin fish schools. Interface Focus 2012,2,693-707.[Cr ossRef] ) ( 8. Filella, A.; Nadal, F. ; Sire, C . ; Kanso, E.; Eloy, C . Model of collective fish behavior with hydrodynamic interactions. Phys. Rev. Lett. 2018,120,198101.[C r o s s Ref] ) ( 9. Lighthill, M.J. Note on the swimming of slender fish. J . Fluid Mech. 1960, 9,305-317. [ C r ossRef] ) ( 10. Blevins, E.; Lauder, G.V. Swimming near the sub s trate: A simple robo t ic model of stingray locom o tion.Bioinspir. Biomim. 2013,8,016005. [ C rossRef] ) ( 11. C Quinn, D.B.; Moored, K.W.; Dewey, P.A.; Smits, A.J. Unsteady propulsion near a solid boundary. J . FluidMech. 2014, 742,152-170. [ CrossRef] ) ( 12. Fernandez-Prats, R.;Raspa, V.; Thiria, B.;Huera-Huarte, F.;Godoy-Diana, R. Large-amplitude undulatoryswimming near a wall. Bioinspir. Biomim. 2015, 1 0 ,016003. [Cr ossRef] [ P u bMe d] ) ( 13. Kurt, M.; Cochran-Carney,J;Zhong, Q.;Mivehchi, A.; Quinn, D.B.; Moored, K. W . Swimming freely near theground l e ads to flow-mediated equilibrium altitudes. J . Fluid Mech. 2019,875, 2125. [C ros s Re f ] ) ( Weihs, D. Hydromechanics of fish schooling. Nature 1973, 241,290-291. [Cros sRef] ) ( Hemelrijk, C.K.; Reid, D.; Hildenbrandt,H.;Padding, J.T. T h e increased efficiency of fish swimming in aschool. Fish Fish. 2015,16,511-521.[ CrossRef] ) ( 16. Daghooghi, M.; Borazjani, I. The hydrodynamic advantages of synchronized swimming in a rectangularpattern. B ioinspir. Biomim. 2015, 10,056018. [C ros s Ref] ) ( 17. Ashraf, I.; Godoy-Diana, R.; Halloy,J.; Collignon, B.; Thiria, B. Synchronization and col l ective swimming patterns in fish (Hemigrammus bleheri). J. R. Soc. Interface 2016, 13,20160734. [ Cross R ef ] ) ( 18. Ashraf, I.; Bradshaw, H.; Ha, T.T.; Halloy,J.; Godoy-Diana, R.;Thiria, B. Sim p le phalanx pattern leads t oenergy saving in c o hesive fish schooling. Pr o c. Na t l. Acad. Sc i . USA 201 7 , 114,9599-9604. [Cros s R e f ] ) ( 19. Collignon, B.; Seguret, A.; Chemtob, Y.; Cazenille,L.; H alloy, J. Col l ective departures and leadership in zebrafish. PLoS ONE 2019, 14, e0216798.[ CrossRef ] ) ( 20. J . Alben, S.; Witt, C.;Baker, T.; Anderson,E.; Lauder,G. Dyn a mics of freely swimming flexible foils. Phys. F luids 2012,24,051901.[C ross R ef ) ( 21. Dewey, P.A.; Boschitsch, B.M.;Mo o red, K.W.; Stone, H.A. ; Smits, A.J. Scaling laws for the thrust productionof flexible pitching panels. J. Fluid Mech. 2013,732,29-46. [ CrossR e f] ) ( 22. Pineirua, M.; Thiria, B.; Godoy-Diana, R. Modelling of an actuated elastic swimmer. J. Fluid Mech. 201 7 ,829,731-750. [C rossRef] ) ( 23. Raspa, V .; R amananarivo, S.; Thiria, B .; Godoy-Diana, R. Vortex-induced drag and the role of aspect ratio in undulatory swimmers. Ph y s. Fluids 2014, 26,041701. [C rossRef] ) ( 24. Li, G.; Kolomenskiy, D.;Liu, H.; Thiria, B.; Godoy-Diana, R. On the energetics and stability of a minimal fishschool. PLoS ONE 2019, 14, e0215265.[C rossRef ) ( 25. Li, G.; Kolomenskiy, D . ; L i u, H.; Thiria, B .; Godoy-Diana, R. On the interference of vorticity and pre s surefields of a minimal fish school. J. Aero Aqua-Bio-Mech. 2019,8, 27-33. [ C rossRef ] ) ( 26. Raspa, V.;Godoy-Diana, R.; Thiria,B. Topology-induced effect in biomimetic propulsive wakes. J. FluidMech. 2013,729,377-387.[C rossRef] ) 27. Weihs, D. Some hydrodynamical aspects of fish schooling. In Swimming and Flying in Nature; Springer:Berlin, Germany, 1975;pp.703-718. 28. Kanso,E.;Newton,P.K. Locomotory Advantages to Flapping Out of Phase. Exp. Mech. 2009,50, 1367-1372.[CrossRefl 29. Huera-Huarte, F.J. Propulsive performance of a pair of pitching foils in staggered configurations. J. FluidsStruct. 2018,81,1-13. [CrossRef] 30. Ramananarivo,S.; Fang, F.;Oza, A.; Zhang, J;Ristroph, L. Flow interactions lead to orderly formations offlapping wings in forward flight. Phys. Rev. Fluids 2016,1,071201. [CrossRef] @ 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open accessCC article distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (http://creativecommons.org/licenses/by/4.0/). In-phase and anti-phase synchronization of neighboring swimmers is examined experimentally using two self-propelled independent flexible foils swimming side-by-side in a water tank. The foils are actuated by pitching oscillations at one extremity—the head of the swimmers—and the flow engendered by their undulations is analyzed using two-dimensional particle image velocimetry in their frontal symmetry plane. Following recent observations on the behavior of real fish, we focus on the comparison between in-phase and anti-phase actuation by fixing all other geometric and kinematic parameters. We show that swimming with a neighbor is beneficial for both synchronizations tested, as compared to swimming alone, with an advantage for the anti-phase synchronization. We show that the advantage of anti-phase synchronization in terms of swimming performance for the two-foil “school” results from the emergence of a periodic coherent jet between the two swimmers.
确定







还剩8页未读,是否继续阅读?

产品配置单


北京欧兰科技发展有限公司为您提供《游泳过程中速度场检测方案(粒子图像测速)》,该方案主要用于其他中速度场检测,参考标准--,《游泳过程中速度场检测方案(粒子图像测速)》用到的仪器有德国LaVision PIV/PLIF粒子成像测速场仪、LaVision DaVis 智能成像软件平台
推荐专场





相关方案
更多
该厂商其他方案
更多