








方案详情
文
When two proteins interact and bind, conformational changes in the proteins, and rearrangement of the solvent in the vicinity of the binding site, result in the absorption or generation of heat. Quantification of this reaction heat by ITC provides a complete thermodynamic description of the binding interaction, the stoichiometry of binding, and the association constant; in addition, if structural information is available, the contributions of specific amino acids mediating the binding event can be identified and their thermodynamic contributions quantified.
方案详情

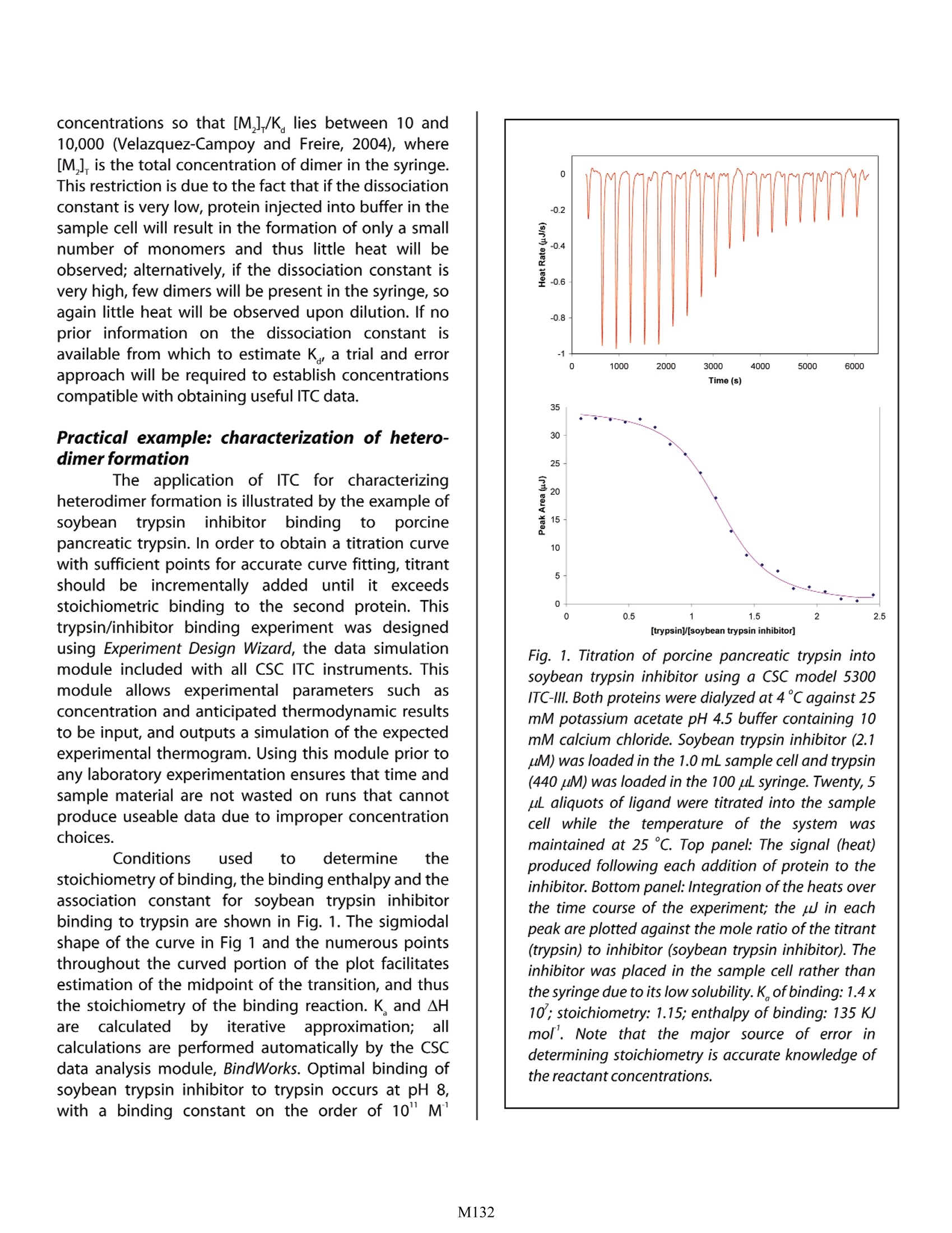
Characterizing Protein-Protein Interactions by ITC Christin T. Choma TA Instruments, 109 Lukens Drive, New Castle, DE 19720,USA roteinss operateasintegral membersS Cofpcomplex networks that precisely regulatephysiological processes through a combinationof signaling pathways and feedback. These regulatoryfunctions,, including signal transduction,i, proteintrafficking, transcription and translation, all rely on theability of proteins to rapidly target and form specificnon-covalent complexesswith other proteinsinresponse too chemicalsignals.Predictingandunderstanding how these interactions occur and arecontrolled (or, in the case of many diseases, how aninteraction misfunctions) iss complicated by thedynamic events controlling recognition and binding.Each interaction results in protein structural changesand involves specific electrostatic and van der Waalsinteractions, the burial of solvent-exposed surfaces,and the breaking and formation of hydrogen bonds. Ithas been estimated that each protein in an organismforms interactions with an average of five otherproteins (Piehler, 2005), and it is known that the sameprotein can form different interactions depending onits location or concentration in the cell (Liddington,2004).. Additionally, aside from such intermolecularinteractions, protein-protein interactions also occurintramolecularly at interfaces between domains andbetween subunits within a protein. This note: examinesthiee application1 ofisothermal titration calorimetry (ITC) for quantitativelymeasuring the thermodynamic properties drivingprotein-protein interactions, and also describes howITc cannbeuused toquantifyprotonation/de-protonation events occurring upon binding. BecauseITC involves the titration of one component intoanother, ITC isgenerally limited to sstudyingbimolecular interactions. Please refer to (CSC'sOverview Note entitled Life Science Applications ofITC' for a general description of the principles behindITC and a summary of the types of biological problemsthat can be addressed by this technique. Analyzing protein-protein interactions Crystal structures of protein complexes showthat the interaction of two protein surfaces requiresgood Ehydrophobic, charge and shape comple-mentarity, so that the resulting interface is as wellpacked as the interior of a pprotein. Given this requirement, coupled with prior knowledge of whichproteins interact (for example, by using yeast two-hybrid systems (Toby and Golemis, 2001), proteinfragment complementation assays (Hu and Kerppola,2003), or a combination of both techniques (Hazbun etal., 2003)), it should in theory be possible to predict theinterfaces through which two proteins will interact,and the mechanism of binding. In practice, however,prediction is complicated by dynamic events occurringin both the protein and in the solvent. The inter- or intramolecular interaction of twoprotein moieties often results in substantial structuralchanges not only in the vicinity of the interactinginterfaces, but also distant from the site of contact(Ladbury and Williams, 2004). This results in alteredhydrogen bonds,electrostatiandhydrophobicinteractions, and in a significant rearrangement ofbound water molecules. Prior to two protein surfacesinteracting, water molecules associated with eachprotein face form organized hydrogen bond networksto polar regions of a protein’s surface, and highly-structured clathrate-like arrangements adjacent tohydrophobic surfaces. When these protein surfacesinteract, many of the organized water molecules arereleased into the bulk solvent, but others remainhighly localized and tightly bound to specific locationson the interacting surfaces. These immobilized watermolecules satisfy otherwise unmet hydrogen bondrequirements to the interface, fill cavities, and mediateinteractions between the opposing interfaces. Becauseof their highly localized and organized structure, thesewaters are believed to be critical to interfacial stabilityand specificity (Raschke, 2006). Indeed, it appears that,at least in some systems, water-mediated interfacialinteractionsaremore specific and stable thaninteractions where water molecules are excluded fromthe protein interface (Ladbury,1996). The interaction of any two protein interfaceswill result in unique protein structural changes and in aunique rearrangement of water molecules distributedbetween the free and bound state. These solventrearrangements result in both entropic and enthalpicchanges to the system: for example, removing waterfrom a non-polar surface requires breaking hydrogenbonds between the solvent molecules (which isenthalpically unfavorable), but the released water molecules are motionally less restricted (which isentropically favorable). Since the thermodynamics ofinteractionsbetweenmolecular interfacesiaredetermined by the number and type ofbonassociating the two surfaces, the balance of theenthalpic and entropiceffects provides athermodynamic fingerprint of the interaction (Ladburyand Chowdhry,1996). ITC isthe mostdirect and quantitativeapproach for characterizing the thermodynamicproperties of protein-protein interactions (Doyle, 1997;Pierce et al., 1999; Jelesarov and Bosshard, 1999).Although other techniques allow the binding affinityand stoichiometry to be determined (Lakey andRaggett, 1998), none are as direct as ITC. For example,spectroscopic studies often require the use of labels,surface plasmon resonance requires immobilization(which can interfere with binding) and analyticalultracentrifugation is time consuming; additionally, theenthalpy of binding cannot be directly measured byany of these approaches, but must be calculated fromthe van’t Hoff relationship. In contrast, by accuratelymeasuring the heat evolved when two proteininterfaces interact, ITC directly provides not only thebinding constant (K,) and stoichiometry (n), but alsothe enthalpy of binding (AH). Additionally, ITC allowsthe change in Gibbs energy of the system, AG, and thechange in entropy, AS, to be obtained from: where R is the universal gas constant and T is theabsolute temperature. Since enthalpic (e.g., hydrogenbonds and van der Waals interactions) and entropic(e.g., solvent mobility) contributions to the stability of acomplex reflect different types of interactions, eachproteinn complex willrespond differentlyittoenvironmental and mutational changes, allowing therelative importance of these interactionss too beassessed (Jelesarovaand Bosshard, 1999).Byconducting the titration in several buffers having thesame pH but different enthalpies of ionization, theenthalpy of ionization and pK of each ionizable groupinvolved in the binding reaction can be estimated (seebelow). Further, by performing the experiment at twotemperatures (T1 and T2), the change in heat capacityof the system,AC, can be obtained from: There is growing evidence that heat capacity effectsreflect not only changes in hydrophobic interactionsbut also in hydrogen bonding and solvation as twoprotein surfaces interact and bind. These solvent-induced heat capacity changes are considerably moresubstantial than previously believed, and can besignificantlyinfluencedby/ protonation events.Jelesarov and Bosshard (1999), Cooper (2005) andPrivalov and Dragan(2006)provide edetaileddiscussions of the physical-chemical basis underlyingheat capacity changes during protein recognition andinteraction. The factors that control protein bindingand complex assembly are critical for designing drugsthat inhibit specific protein-protein interactions (Zhaoand Chmielewski, 2005), and have heightened theimportance of obtaining a complete thermodynamicdescription of protein folding and protein-proteininteraction processes. Practicall considerations:Typesopffpirotein-protein interactions compatible with ITC There are three distinct categories of protein-protein interactions: domain-domain, heteromeric,and homomeric. In domain-domain interactions, twoindependently-folded domains form an ;i interface,often involving residues widely separated in sequence.Because domain-domain interactions occur within thesame polypeptide chain, they cannot be studied by ITCexperiments ((wwhhiicchh involve the titration of onecomponent into another) unless protein fragments areused. Instead, domain-domain interactions are oftenprobed by mutating key residues thought to beimportant to domain interactions, and determiningthe stability of the mutants by differential scanningcalorimetry (DSC). Please refer to CSC's applicationnote entitled: Characterizing protein stability by DSC'for a description of the general approach. ITC is ideally suited to studying heterodimericinteractions (Velazquez-Campoy et al.,2004).Asolution containing one protein (in the ITC syringe) isinjected incrementally into a solution of the secondprotein (in the calorimetric cell). The binding orassociation of the two proteins with each stepwiseaddition of titrant releases heat proportional theconcentration of the complex following injection i: where q is the heat released by the ith injection, V isthe volume of the cell (a constant), AH is the enthalpyof association, and [M,M,]. is the concentration of thecomplex after the ith injection. Titrant (M,) is added tothe second protein(M,) until it is saturated and nofurther binding is observed. Since the concentrationsof the two proteins are known throughout thetitration, nonlinear regression analysis of q, allowscalculation of AH, the binding constant(K=[M,M,]/[M,][M,]),and of AS. ITC is also suited for studying the dissociationof homodimers. The dimeric protein is titrated intobuffer in the calorimetric cell, where dilution of theprotein leads to dissociation into its constituentsubunits (Velazquez-Campoyet al.,2004).Thedissociation constant K. determines the distribution ofthe subunits between the monomeric (M) and dimeric(M,) states (K.=[M]/[M,]). Since the heat associatedwith each injection of protein (q) is proportional to theconcentration of monomer after the ith injection, theenthalpy due to the dissociation of the protein intomonomers (AH) can be determined by: where V is again the volume of the calorimetric cell, v isthe injection volume, [M], is the concentration ofprotein in the cell (on a per monomer basis),and F, acorrection factor, is the fraction of monomer present inthe dimer solution in the syringe (since dimers alwaysexist in equilibrium with monomers). As the titrationproceeds and the monomer concentration in the cellincreases, the dissociation reactionnbecomesincreasingly lessfavorable: until finally, furtherinjections of dimer produce no significant heat. Thedissociation constant, the enthalpy due to dissociationand the entropy can then be determined by nonlinearregression analysis. Concentration ranges compatible with ITC Since equilibriuml processesiiinvolve thereversible interconversion of two components (such astwo proteins to a protein complex, or a dimer tomonomers), experimentssrmust be conductedat appropriate concentrations so that a change in theconcentration of one component leads to ameasurable change in the concentration of the secondcomponent. For binding experiments in which twoproteins interact, concentrations should be chosen sothat during approximately the first third of thetitration, essentially all the titrant bindss to themacromolecule in the cell, and during the last third ofthe titration,theermacromoleculeis essentiallysaturated with titrant and little or no further bindingOccurs.When heatt Vs.[M,]/[M,] isplotted, thecurvature in the center of the plot (between these twoextremes) allowsiaccurate determination of theenthalpy and the association constant for the bindingreaction. These conditions are generally achieved ifconcentrations are chosen so that: where [M]., istthe total concentrationoffthemacromolecule in the reaction cell. Binding constantsin the order of 10 - 10°M’ can be accuratelycalculated; binding constants for systems with tighterbinding can often be estimated by performing thetitration at suboptimal pH or temperature, and thenextrapolating K to the desired conditions (Doyle et al.,1995; Doyle and Hensley, 1998; Velazuqez-Campoyand Freire, 2005). Alternatively, accuratedeterminations of very high or low binding constantscan)be obtainedthrough competitive tbindingexperiments; see CSC's application note:entitled"Characterizing binding interactions by ITC' for a fulldescription of this approach. If an accurate determination of the enthalpy ofbinding is of primary importance, the 'titrant’ proteinshould be injected into a large excess of the 'binding'protein in the sample cell. Although neither thebinding constant or the stoichiometry of binding canbe determined from this experiment (since nosaturation of the binding site will occur), the bindingheat can be very accurately measured followingsubtraction of a blank run designed to compensate fordilution effects, buffer mismatch, etc. (Jelesarov andBosshard,1999;O'Brien et al., 2001). The dissociation of dimeric proteinss withdissociation constants in the range 10-10'M can bedetermined by ITC if the experiments are conducted at concentrations so that [M,]/K lies between 10 and10,000 (Velazquez-Campoy and Freire, 2004), where[M,] is the total concentration of dimer in the syringe.This restriction is due to the fact that if the dissociationconstant is very low, protein injected into buffer in thesample cell will result in the formation of only a smallnumber of monomers and thus little heat will beobserved; alternatively, if the dissociation constant isvery high, few dimers will be present in the syringe, soagain little heat will be observed upon dilution. If noprior information on the dissociation constant isavailable from which to estimate K., a trial and errorapproach will be required to establish concentrationscompatible with obtaining useful ITC data. Practical example: characterization of hetero-dimer formation The application of ITC for characterizingheterodimer formation is illustrated by the example ofsoybean trypsin iinhibitor bindingto)porcinepancreatic trypsin. In order to obtain a titration curvewith sufficient points for accurate curve fitting, titrantshould be incrementally added until it exceedsstoichiometric binding to the second protein. Thistrypsin/inhibitor binding experiment was designedusing Experiment Design Wizard, the data simulationmodule included with all CSC ITC instruments. Thismodule allows experimentalparameters such asconcentration and anticipated thermodynamic resultsto be input, and outputs a simulation of the expectedexperimental thermogram. Using this module prior toany laboratory experimentation ensures that time andsample material are not wasted on runs that cannotproduce useable data due to improper concentrationchoices. Conditions used to determine thestoichiometry of binding, the binding enthalpy and theassociation constant for soybean trypsin inhibitorbinding to trypsin are shown in Fig.1. The sigmiodalshape of the curve in Fig 1 and the numerous pointsthroughout the curved portion of the plot facilitatesestimation of the midpoint of the transition, and thusthe stoichiometry of the binding reaction. K and AHaree calculatedbbyiterative approximation; allcalculations are performed automatically by the CSCdata analysis module, BindWorks.Optimal binding ofsoybean trypsin inhibitor to trypsin occurs at pH 8,with a binding constant on the order of 10" M' Fig. 1. Titration of porcine pancreatic trypsin intosoybean trypsin inhibitor using a CSC model 5300ITC-III. Both proteins were dialyzed at 4℃ against 25mM potassium acetate pH 4.5 buffer containing 10mM calcium chloride. Soybean trypsin inhibitor (2.1uM) was loaded in the 1.0 mL sample cell and trypsin(440 uM) was loaded in the 100 ul syringe.Twenty, 5uL aliquots of ligand were titrated into the samplecell while the temperature of the systeml WaSmaintained at 25℃. Top panel: The signal (heat)produced following each addition of protein to theinhibitor. Bottom panel: Integration of the heats overthe time course of the experiment; the pJ in eachpeak are plotted against the mole ratio of the titrant(trypsin) to inhibitor (soybean trypsin inhibitor). Theinhibitor was placed in the sample cell rather thanthe syringe due to its low solubility. K,of binding: 1.4x10 ; stoichiometry: 1.15; enthalpy of binding:135 KJmof. Note that the major source of error indetermining stoichiometry is accurate knowledge ofthe reactant concentrations. (Velazquez-Campoy et al., 2004). The present titrationwas conducted at 25℃, pH 4.5 in order to decrease theaffinity to a level compatible with ITC (range 10”-10°M'). Under these conditions, the proteins; bindendothermically with an enthalpy of 135 KJ mol and aK of 1.4 x 10'. It is possible to estimate the heatcapacity change upon association by repeating thetitration at several temperatures and plotting theenthalpy of association vs. temperature; the slope ofthe plot corresponds to the change in heat capacitydue to association. A positive value is indicative ofprotein surface being exposed to solvent when twoproteins interact (which is more likely to occur whentwo proteins dissociate rather then bind), whereas anegative value reflects the dehydration of proteinsurfaces (Privalov and Dragan, 2006). Protonation effects When two protein interfaces bind, the pkvalues of ionizable groups at the interfaces can bealtered due to solvation or electrostatic changestriggeredbythe: bindingevent,resultinginprotonation or deprotonation of these residues. If thisoccurs, the association constant will be pH-dependent,and the binding enthalpy will be dependent on theionization enthalpy of the buffer (AH). A series of ITCexperimentsSccan(establishwhetheraaprotona-tion/deprotonation reaction is occurring and if so, howmany protons are involved. If independent structuralinformation about the interacting residues in thecomplex isbavailable, individual contributions ofspecific amino acids to the binding reaction can bedetermined, allowing a thermodynamic mapping ofthe binding interface (Gomez and Freire, 1995; Frischet al.,1997;Crnogorac et al., 2001) The ITC titrations are performed in buffers withdifferentionizationenthalpies. Theemeasuredenthalpy, AHa is equal to the enthalpy of binding(AHind which is independent of the buffer used, but ispH dependent), to n,the number of protons adsorbedor released by the binding reaction, and to theionization enthalpy of the buffer: The buffer ionization enthalpy can be obtained fromtables (Christensen et al., 1976), measured (Jelesarov and Bosshard,1994) or obtained from the papers citedbelow. A plot of AHvs. the buffer ionization enthalpywill provide a straight line from which n can beestimated from the slope and AH from the interceptwith the y axis. If the slope is zero then there is no nettransfer of protons, if n. is positive then protons aretransferred from the buffer solvent to the proteincomplex, and if n, is negative, then protons aretransferred from the complex to the solvent. Theenthalpy of ionization and the pK of each ionizablegroup involved in the protonation/deprotonationreaction can be calculated using the approachdeveloped by Baker and Murphy (1996 and 1997) andGomez and Freire (1995). This approach has been usedto quantify the binding energetics of closely-relatedinhibitors (Parker et al., 1999), the displacement ofligands (Ciaccio et al., 2004) and competing metals(Christensen et al.,2003), and the identification ofcharged groups involved in binding drugs to proteinsanrand DNA (Velazquez-Campoy et al., 2000; Kaul et al.,2003; Lobo et al., 2003; Barbieri and Pilch, 2006;Nguyen et al., 2006). Summary Protein-protein interactions mediateessentially all physiological processes and areimplicated in a large number of disease states. There istherefore increasing interest in understanding thethermodynamics driving specific binding interactions,with the intent that the selective control or disruptionof these interactions could help elucidate fundamentalbiological mechanisms. ITC is a straightforward andcompletely general aapppprrooaacchh for determiningiiacomplete thermodynamic description of the bindinginteractions between two proteins, as well as thestoichiometry of binding and the association constant.In addition, if structural information about the bindinginterface is available, the contributions of specificamino acids mediating the binding event can beidentified and their thermodynamic contributionsquantified. References (Preference has been given to current references. Citation doesnot imply that a paper is necessarily the original reference to astudy.) Baker, B. M. and K. P. Murphy. (1996) Evaluation oflinked1 protonation effects in protein bindingreactions using isothermal titration calorimetry.Biophys.J.71,2049-2055. Baker, B. M. and K. P. Murphy. (1997) Dissecting theenergetics of a protein-protein interaction: thebinding of ovomucoid third domain to elastase.J.Biol. Chem.268,557-569. Barbieri, C. M. and D. S. Pilch. (2006) Completethermodynamic characterization of the multipleprotonation equilibria of the aminoglycosideantibiotic paromomycin: a calorimetric and naturalabundance N NMR study. Biophys. J. 90, 1338-1349. Christensen, J. J., L. D. Hansen and R. M. Izatt. (1976).Handbook of proton ionization heats and relatedthermodynamic quantities. Wiley, New York. Christensen, T., D. M. Goodin, J. E. King and E. J. Toone.2003 Additivity/ and thephysicallbasis5otmultivalency effects: a thermodynamicinvestigation of the calcium EDTA interaction. JACS125,7357-7366. ( Ciaccio, C. et a l . (2004) Proto n linkag e for CO binding and r edox properties of b ovine l a ctoperoxidase. Biophys.J.86,448-454. ) ( Crnogorac, M. M., G. M. Ullmann a nd N. M. Kostic.(2001) E ffects of pH on protein association: Modification of the p rotein-linkage model a n d experimental verification of the modified model in the case of cytochrome c and p l astocyanin. JACS 123,10789-10798. ) Doyle, M. L. (1997) Characterization of bindinginteractions by isothermal titration calorimetry.Curr. Opin. Biotechnol.8,31-35. ( Doyle, M . L ., G. L ouie, P . R . Dal M onte and T. K. Sokoloski. (1995) Tight binding affinities determined from n t thermodynamic linkage e t to protons by t i tration c alorimetry. M eth. Enzymol. 259,183-194. ) Doyle, M. L. and P. Hensley. (1998) Tight ligand bindingaffinities determined from thermodynamic linkageto temperature by titration calorimetry. Meth.Enzymol.295,88-99. Frisch, C., G. Schreiber, C. M. Johnson and A. R. Fersht.(1997) Thermodynamics of the interaction ofBarnase and Barstar: changes in free energy versuschanges in enthalpy on mutation.J. Biol. Chem.267,696-706. Gomez, and E. Freire. (1995) Thermodynamicmapping of the inhibitor site of the asparticprotease endothiapepsin.J.Mol. Biol. 252, 337-350 Hazbun, T. R. et al. (2003) Assigning function to yeastproteins by integration of technologies.Mol. Cell.12,1353-1365. Hu, C. D. and T. K. Kerppola. (2003) Simultaneousvisualization of multiple protein interactions inliving cells usingmulticolorfluorescencecomplementation analysis. Nat. Biotechnol. 21,539-545. Jelesarov,l and H. R. Bosshard. (1994) Thermodynamicsofferredoxinbindingyttoferredoxin:NADP+reductase and the role of water at the complexinterface. Biochemistry 33,13321-13328. ( Jelesarov, l . and H. R. Bosshard. (1999) Isothermal titratio n calorimetry and differential s scanning calorimetry as complementary tools to investigatethe energetics of biomolecular recognition. J. Mol.Recognit. 12,3-10. ) Kaul, M., C. M. Barbieri, J. E. Kerrigan and D. S. Pilch.(2003) Coupling of drug protonation to the specificbinding of aminoglycosides to the A site of 16 SrRNA: elucidation of the number of drug aminogroups involved and their identities. J. Mol. Biol.326, 1373-1387. Ladbury, J. E. (1996) Just add water! The effect of wateron the specificity of protein-ligand binding sitesand its potential application to drug design. Curr.Opin. Struct. Biol. 3,973-980. Ladbury, J. E. and B. Z. Chowdhry.(1996) Sensing theheat: the application of isothermaltitrationcalorimety tothermoydynamicstudiesofbiomolecular interactions. Chemistry & Biology 3,791-801. Ladbury, J. E. and M. A. Williams. (2004) The extendedinterface:measuringnon-local effects inbiomolecular interactions. Curr. Opin. Struct. Biol.14,562-569. Lakey, J. H. and E. M. Raggett. (1998) Measuringprotein-protein interactions. Curr. Opin. Struct. Biol.8,119-123. Liddington, R. C. (2004) Structural basis of protein-protein interactions. In H. Fu (Ed.) Methods inMolecular Biology,vol.261:FProtein-proteininteractions: methodsandprotocols.pp.3-13,Humana Press,Totowa,NJ. Lobo, B. A., G. S. Koe, J. G. Koe and C. R. Middaugh.(2003) Thermodynamic analysis of binding andprotonation in DOTAP:DOPE (1:1):DNA complexesusing isothermal titration calorimetry. Biophys. J.104,67-78. Nguyen, B., J. Stanek and W. D. Wilson. (2006) Binding-linked protonation of a DNA minor-groove agent.Biophys.J.90,1319-1328. ( O'Brien, R. , J . E . Ladbury and B . Z . Chowdry. (2001) Isothermal titration calorimetry of biomolecules. p.263-286. In S. E. Harding and B. Z . Chowdry (Eds.) Protein-Ligand Interactions : hydrodynamics and calorimetry.Oxford University Press, Oxford. ) ( Parker, M. H. et al. (1999) Analysis of the binding of hydroxamic a c id a n d carboxylic a c id i n hibitors t o the stromelysin- 1 1 ( matrix m etalloproteinase-3) catalytic domain 1 by isothermal titration calorimetry. Biochemistry 38, 1 3592-13601. ) ( Piehler, J. (2005) New methodologies f or measuring protein interactions i n vivo and in vitro. Curr. O pin. Struct.Biol. 15,4-14. ) ( Pierce, M. M., C. S. Raman and B. T. Nall. (1999) Isothermal titration calorimetry of protein-protein interactions.Methods 19,213-221. ) Privalov, P. L. and A. I. Dragan. (2006) Microcalorimetryof biological macromolecules. Biophys. Chem. 122,158-169. Raschke, T. M. (2006) Water structure and interactionswith protein surfaces. Curr. Opin. Struct. Biol. 16,152-159. Toby, G. G. and E. A. Golemis.(2001) Using the yeastinteraction trap and other two-hybrid-basedapproaches to study protein-protein interactions.Methods 24,201-217. ( Velazquez-Campoy, A., L . . Luque,M . . J. Todd, M.Milutinovic h, Y. Kiso and E. F Freire . (2000) Thermodynamic dissection OT the binding energetics of KNT-272, a potent HIV-1 protease inhibitor.Prot. Sci. 9 ,1801-1809. ) Velazquez-Campoy, A., S. A. Leavitt and E. Freire.(2004)Characterization of protein-protein interactions byisothermal titration calorimetry. In H. Fu (Ed.)Methods in Molecular Biology, vol. 261: Protein-protein interactions: methods and protocols. pp.35-54,Humana Press, Totowa, NJ. ( Velazquez-Campoy a n d E. F r eire. ( 2005). ITC in the post-genomic C era...! Priceless. B iophysical Chemistry 115,115-124. ) ( Zhao, L . and J . Chmielewski. ( 2005) I nhibiting protein- protein i nteractions using d esigned m o lecules. Curr. Opin. Struct. Biol.15,31-34. ) M When two proteins interact and bind, conformational changes in the proteins, and rearrangement of the solvent in the vicinity of the binding site, result in the absorption or generation of heat. Quantification of this reaction heat by ITC provides a complete thermodynamic description of the binding interaction, the stoichiometry of binding, and the association constant; in addition, if structural information is available, the contributions of specific amino acids mediating the binding event can be identified and their thermodynamic contributions quantified.
确定






还剩6页未读,是否继续阅读?

产品配置单

TA仪器为您提供《蛋白质中交互作用检测方案(差示扫描量热)》,该方案主要用于其他中交互作用检测,参考标准--,《蛋白质中交互作用检测方案(差示扫描量热)》用到的仪器有TA仪器+Affinity ITC+等温滴定微量热仪
推荐专场






相关方案
更多
该厂商其他方案
更多