








过热蒸汽对熟制小龙虾优势腐败菌的杀菌动力学及其机理
方案详情

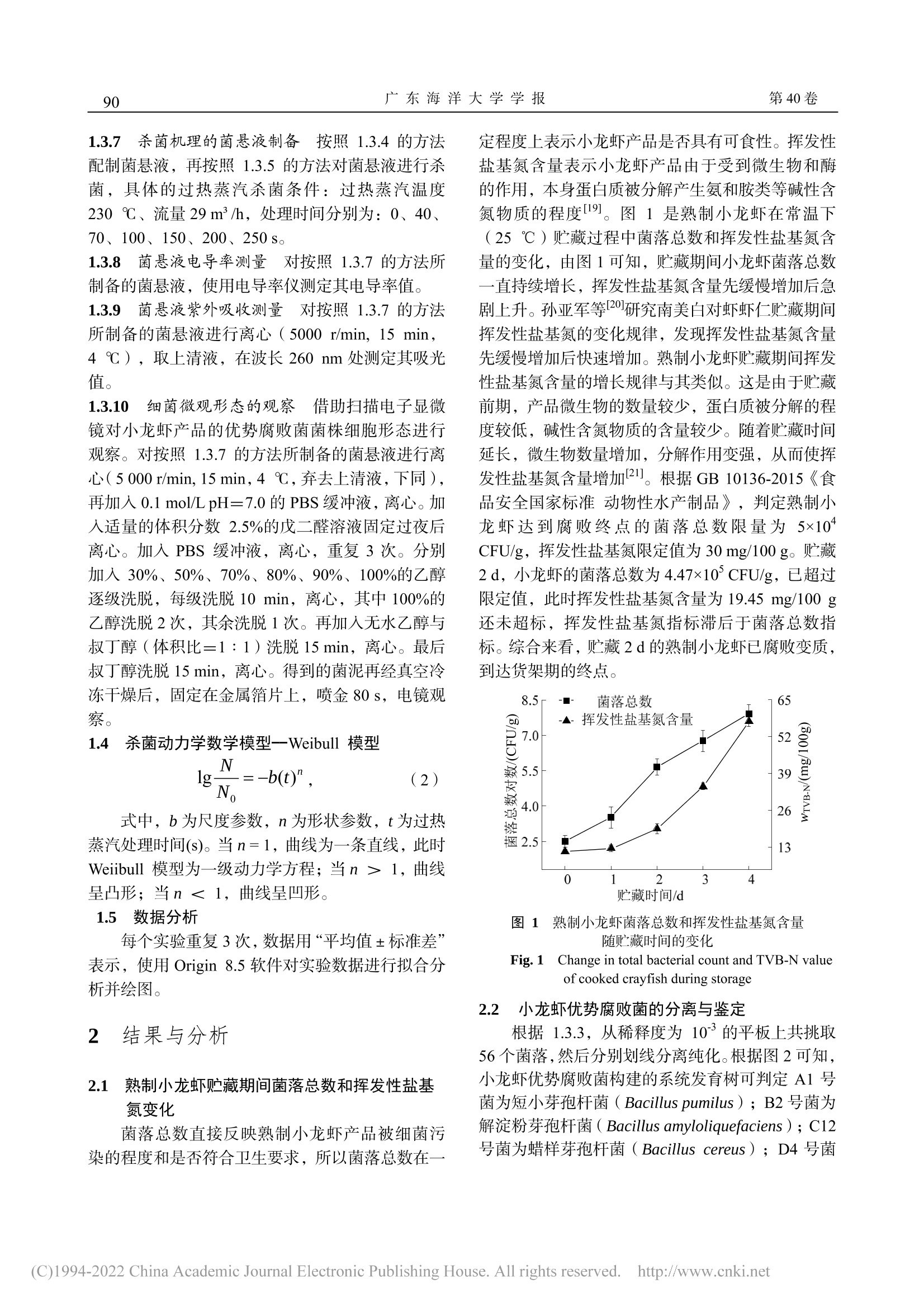
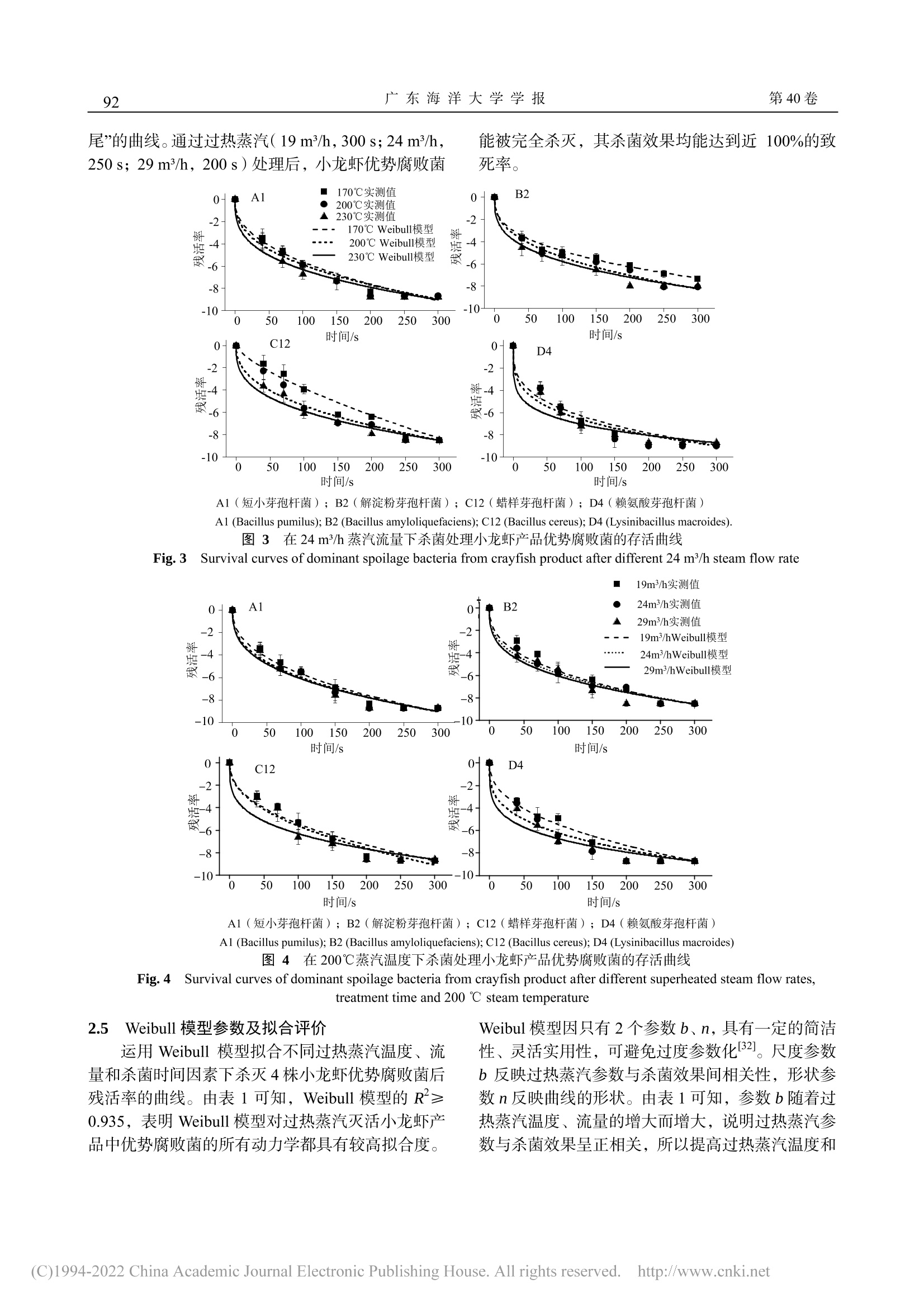
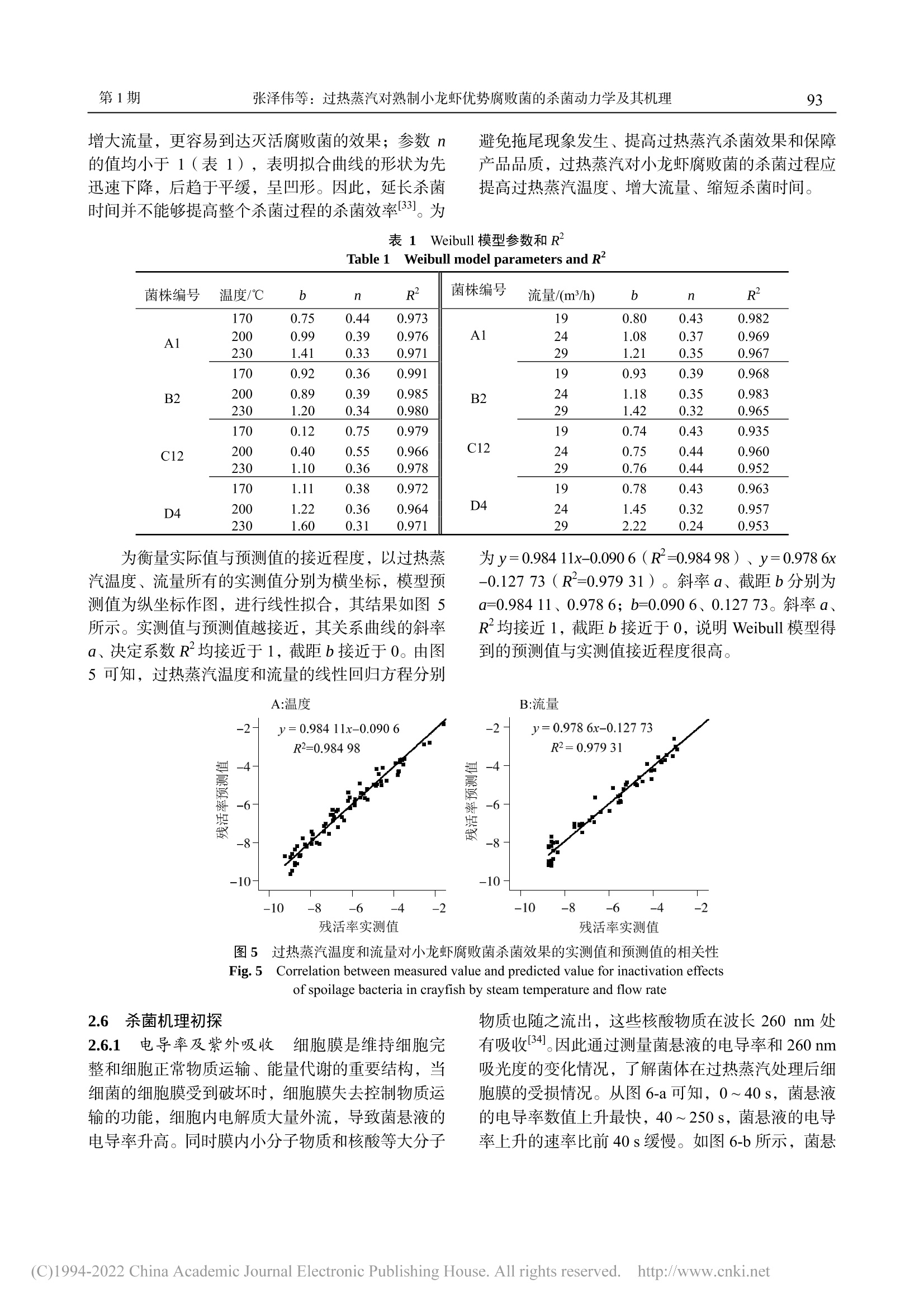
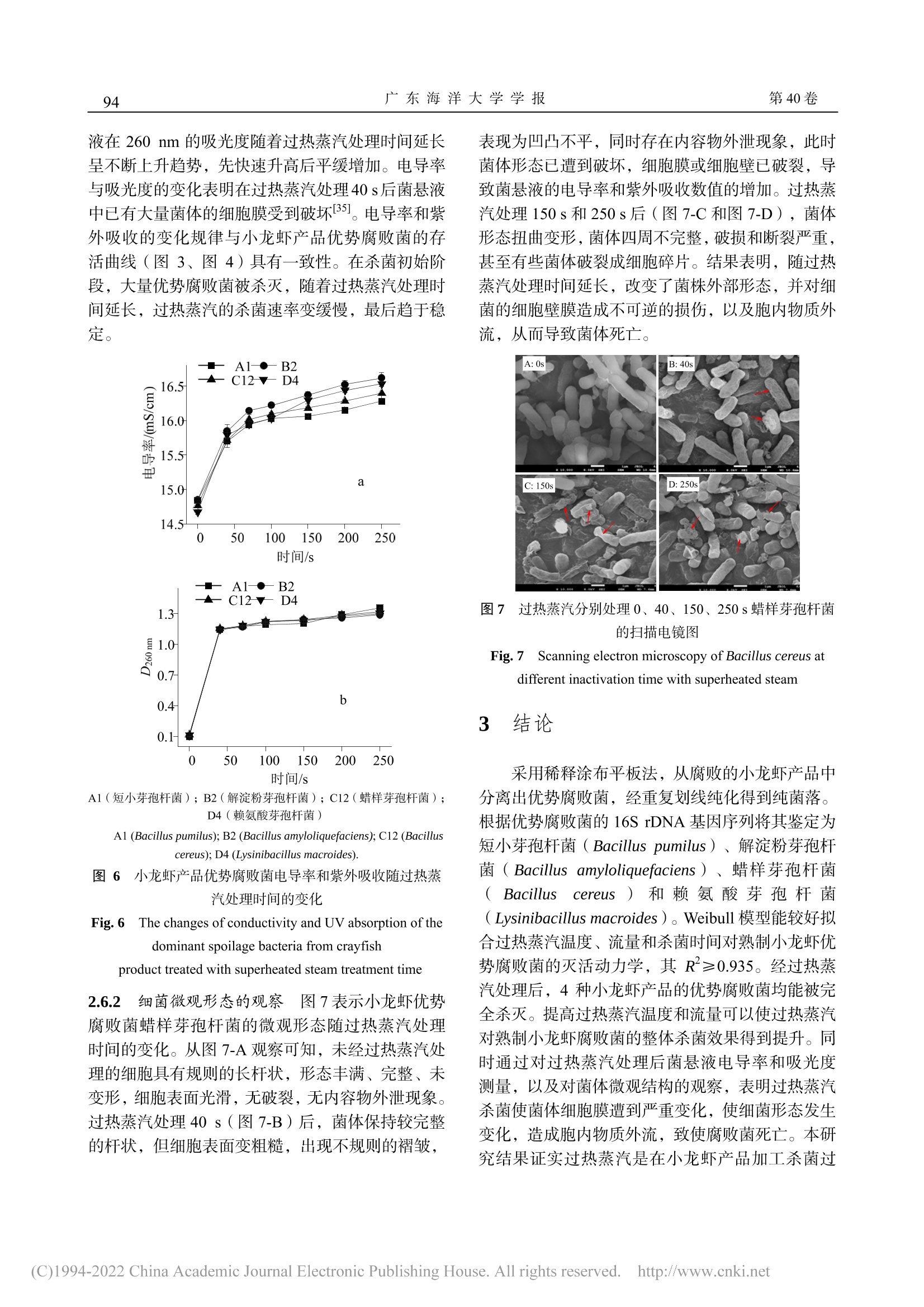
过热蒸汽对熟制小龙虾优势腐败菌的杀菌动力学及其机理广东海洋大学学报Journal of Guangdong Ocean University第 40卷 第 1期2020年 1月Vol.40 No.1Jan. 2020 第 40卷广东海洋大学学报88 张泽伟,段伟文,陈铭,等 .过热蒸汽对熟制小龙虾优势腐败菌的杀菌动力学及其机理 [J].广东海洋大学学报,2020,40(1):87-96. 过热蒸汽对熟制小龙虾优势腐败菌的杀菌 动力学及其机理 张泽伟 1,段伟文 1,陈 铭 1,邓楚津 1,刘书成 1,2,3,4,吉宏武 1,2,3,4 (1.广东海洋大学食品科技学院 / 2.广东省水产品加工与安全重点实验室 / 3.广东省海洋食品工程技术研究中心 /4.水产品深加工广东普通高等学校重点实验室,广东湛江 524088) 摘 要:【目的】研究过热蒸汽杀菌对熟制小龙虾优势腐败菌的杀菌效果。【方法】从腐败熟制小龙虾中分离腐 败菌,根据 16S rDNA 基因序列信息鉴定其种属,研究过热蒸汽温度、流量和杀菌时间等的处理参数对小龙虾优 势腐败菌的杀菌效果,并利用 Weibull 模型拟合其杀菌动力学过程。通过研究过热蒸汽处理对优势腐败菌菌悬液的 电导率和紫外吸收的影响,以及通过扫描电子显微镜对菌体的微观结构进行观察,初步探讨其杀菌机理。【结果】熟制小龙虾优势腐败菌为短小芽孢杆菌(Bacillus pumilus )、解淀粉芽孢杆菌(Bacillus amyloliquefaciens )、蜡样 芽孢杆菌(Bacillus cereus )和赖氨酸芽孢杆菌(Lysinibacillus macroides )。在过热蒸汽杀菌初期阶段,优势腐败 菌的残活率迅速下降,随着杀菌时间延长,残活率下降速率变得缓慢,最后趋于稳定,说明过热蒸汽能完全杀灭 小龙虾优势腐败菌。Weibull 模型能较好拟合过热蒸汽对 4种优势腐败菌的杀菌动力学,其 R 2≥0.935。Weibull 模 型得到的预测值与实测值接近程度较高。通过对模型的分析,提高过热蒸汽温度和增大流量,更容易到达灭活腐 败菌的效果。过热蒸汽杀菌后,菌悬液的电导率值和吸光度迅速上升,之后趋于平缓;通过对微观结构的观察,发现杀菌后腐败菌的细胞膜完整性遭到严重破坏。【结论】过热蒸汽杀菌技术应用于小龙虾产品加工具有一定的 可行性。 关键词:过热蒸汽 ;杀菌动力学 ;小龙虾优势腐败菌 ;杀菌机理 中图分类号 :TS201.3;TS254.4文献标志码 :A 文章编号 :1673-9159(2020)01-0087-10 doi :10.3969/j.issn.1673-9159.2020.01.012 Inactivation Kinetics and Mechanisms of Dominant Spoilage Bacteria in Cooked Crayfish (Procambarus clarkii) by Superheated Steam ZHANG Ze-wei1 , DUAN Wei-wen1, CHEN Ming1, DENG Chu-jin1, LIU Shu-cheng1,2,3,4, JI Hong-wu1,2,3,4 (1. College of Food Science and Technology , Guangdong Ocean University , Zhanjiang 524088, China ;2. Guangdong Provincial Key Laboratory of Aquatic Product Processing and Safety, Zhanjiang 524088 , China ; 3. Guangdong Provincial Engineering Technology Research Center of Marine Food , Zhanjiang 524088, China ; 4. Key Laboratory of Advanced Processing of Aquatic Product of Guangdong Higher Education Institution , Zhanjiang 524088, China ) 收稿日期 :2019-08-22 基金项目 :国家虾蟹产业技术体系建设专项( CARS-48) 第一作者 :张泽伟(1994-),男,硕士研究生,研究方向为水产品加工及贮藏工程。Email :18320422276@163.com 通信作者 :吉宏武(1962-),男,博士,教授,研究方向为水产品加工及贮藏工程。Email :jihw62318@163.com Abstract : 【Objective 】The aim of this work was to study the bactericidal effect of superheated steam treatment on dominant spoilage bacteria from cooked crayfish. 【Method 】The dominant spoilage bacteria were isolated from the rotten cooked crayfish samples and identified by the 16S rDNA gene sequences information. The effects of superheated steam treatment parameters such as temperature, flow rate and time on the dominant spoilage bacteria isolated were investigated. Inactivation kinetics of the dominantspoilagebacteriabysuperheatedsteamwasfittedbyusingtheWeibullmodel.The sterilization mechanism of superheated steam was explained by analyzing the values of conductivity and ultraviolet absorption, and observing the microbial morphology of the bacteria. 【Result 】The results showed that the dominant spoilage bacteria of the cooked crayfish were identified as Bacillus pumilus , Bacillusamyloliquefaciens ,Bacilluscereus and Lysinibacillusmacroides .Intheinitialstageof superheated steam sterilization, the survival rate of dominant spoilage bacteria decreased rapidly, and with the prolongation of sterilization time, the decline rate became slow, and finally tended to be stable. It indicated that superheated steam could completely inactivate the dominant putrefactive bacteria of crayfish. Inactivation kinetics curves of 4 colonies of the dominant spoilage bacteria by superheated steam were well fitted by the Weibull model ,with R2≥0.935. The predicted value of the Weibull model is close to the measured value. According to the analysis of the model, increasing of superheated steam temperature and flow rate and shortening of the sterilization time could achieve better inactivation effect onspoilagebacteriafromcrayfish.Aftersuperheatedsteamtreatment,theconductivityandUV absorbance of bacterial suspension increased rapidly, and the observation of the microbial morphology of the bacteria showed that the bacterial cell membrane was seriously damaged, resulting in the death of the bacteria, which may be one of the main reasons to reach inactivation. 【Conclusion 】Superheated steam technology was a promising sterilization technology during crayfish products processing. Keywords:superheatedsteam;inactivationkinetics;dominantspoilagebacteriafromcrayfish; sterilizing mechanism 小龙虾,学名克氏原螯虾( Procambarus clarkii ),自 1929年引入我国后,被大范围推广养 殖。2016年,我国已是全世界最大小龙虾生产国,其加工产业快速发展的同时也遇到了诸多问题 [1-2]。与其他冷藏水产品相比,小龙虾含有较多自由氨基 酸,致使其更容易遭受微生物作用而腐败变质,货 架期较短,成为限制小龙虾产业进一步发展的主要 障碍 [3]。如何有效地杀灭小龙虾产品中腐败菌,前 人已有相关杀菌方式的研究报道。其中巴氏灭菌能 较大程度保持小龙虾产品灭菌后品质,但杀菌效果 不理想。高温高压灭菌能较好杀灭微生物,但对产 品风味与口感等影响较大 [4]。因此选择合适杀菌方 式是小龙虾产业急需解决的问题。 过热蒸汽是指在恒定的压力下对饱和蒸汽继 续加热,使其温度升高到相应沸点以上的高温水蒸 气 [5]。与传统热处理方式相比,过热蒸汽的传热方 式除对流传热和辐射传热,还有冷凝传热;在杀菌 过程中蒸汽在物料表面冷凝,释放大量的热,使物 料快速升温同时大幅度杀灭微生物,缩短杀菌时 间,保障产品品质。且在杀菌过程中过热蒸汽提供 低氧环境,避免氧化反应,降低有害物生成。近些 年过热蒸汽杀菌作为一种绿色热加工技术而备受 关注 [6]。目前,研究已发现过热蒸汽对大肠杆菌 (Escherichia coli )[7]、嗜热芽孢杆菌(Geobacillus stearothermophilus )[8]、沙门氏菌(Salmonella )[9]等微生物具有良好的杀菌效果。然而,过热蒸汽杀 菌技术对小龙虾产品优势腐败菌杀灭效果的研究 尚未有文献报道。 微生物杀菌动力学研究是杀菌技术运用的关 键理论之一,它可以定量评价杀菌的效果、提供合 适的杀菌设计标准,对杀菌技术的实际应用具有理 论指导意义 [10]。目前,已研究多种杀菌技术对不同 微生物的杀菌动力学,例如超声辅助超临界 CO2杀 灭大肠杆菌(E. coli )和酿酒酵母(Saccharomyces cerevisiae )的动力学 [11],微波和巴氏灭菌杀灭大肠 杆菌 O157:H7(Escherichia coli O157:H7)和李 斯特菌(Listeriamonocytogenes )的动力学 [12]和过 热蒸汽杀灭沙门氏菌和蜡样芽孢杆菌的杀菌动力 学 [13]。然而,过热蒸汽对小龙虾优势腐败菌的杀菌 动力学还未见有研究报道。 本研究以从腐败小龙虾产品中分离的优势腐 败菌为研究对象,研究不同条件下过热蒸汽温度、流量和处理时间对小龙虾产品优势腐败菌的杀菌 效果及动力学,并初步探究其杀菌机理,以期获得 过热蒸汽杀灭小龙虾产品优势腐败菌的最佳条件,为指导过热蒸汽在小龙虾产品加工中的实际应用 提供理论依据。 1材料与方法 1.1 材料与试剂 鲜活小龙虾(Procambarus clarkii ),30 ~ 36尾 /kg ,购于广东省湛江市霞山区东风海鲜市场;营养肉汤 (NB)、PCA 平板计数琼脂、革兰氏染色试剂盒,北 京陆桥技术股份有限公司;Taq DNA 聚合酶、PCR 引物,生工生物工程 (上海 )股份有限公司;基因组 DNA 提取试剂盒,天根生化科技有限公司;体积分 数 25%戊二醛、无水乙醇、叔丁醇,均为分析纯,广东光华科技股份有限公司。 1.2 主要实验仪器和设备 JSM-7610F 扫描电子显微镜,日本电子株式会 社;SW-CJ-FD 洁净双人工作台,苏州净化设备有 限公司;LDZX-50KBS 立式压力蒸汽灭菌器,上海 申安医疗器械厂;Hema 9600 PCR 基因扩增仪,珠 海黑马医疗仪器有限公司;SLI-700埃朗生化培养 箱,东京理化器株式会社;HZQ-X100全温振荡培 养箱,苏州培英实验设备有限公司;FDU-1100真空 冷冻干燥设备,东京理化器械株式会社;Aglient Cary 60紫外 -可见光光度计,美国安捷伦科技公司; DDSJ-308A 电导率仪,上海仪电科学仪器股份有限 公司;过热蒸汽处理系统,自制 [14]。 1.3 实验方法 1.3.1 样品制备 新鲜小龙虾 →超声清洗 →沥干 →油炸(170 ℃油炸 3 min )→煮制(含 5 g/100 mL 食盐的沸水中煮 10 min )→冷却 →真空包装 →25 ℃下贮藏。 1.3.2 贮藏期间菌落总数与挥发性盐基氮含量测 定 每天测定熟制小龙虾的菌落总数与挥发性盐 基氮含量。菌落总数测定方法参照 GB 4789.2-2016 《食品安全国家标准食品微生物学检验菌落总 数测定》;挥发性盐基氮采用自动凯氏定氮仪法测 定,参照 GB 5009.228-2016《食品安全国家标准食 品中挥发性盐基氮的测定》。 1.3.3 腐败菌的分离纯化与鉴定 选取贮藏终点 的小龙虾样品,在无菌超净台中将小龙虾去头、去 壳,称取 25g 虾肉,用无菌剪刀剪碎后放入盛有 225 mL 无菌生理盐水的锥形瓶中,均质。按 10倍 系列稀释,稀释至 10-5。取 100 μL 菌悬液均匀涂抹 在营养琼脂上,在 37℃下,恒温培养 48 h 左右。挑选一个合适的平板(菌落数约 30 ~100 CFU )计 数,并挑起平板上全部可见菌落,进行编号,连续 进行 3次划线分离。用体积分数为 20%甘油-80 ℃保存,以备后用。根据革兰氏染色、细菌形态和菌 落特征将获得的菌株分组。将所有纯菌株进行 16S rDNA 的 PCR 扩增,其产物送往生工生物工程(上 海)股份有限公司测序 [15]。将测序结果在 NCBI 网 站上进行 BLAST 系列分析,选取相似性 99%以上 菌株的基因序列,利用 MEGA5.1软件进行邻近法 (Neighbor-Joining )构建系统发育树。 1.3.4 菌悬液的配制 用接种环挑取一环小龙虾 的优势腐败菌接入无菌的营养肉汤(NB )中,180r/min 、37℃振荡培养 24 h ,连续重复 3次 [16]。将 培养好的细菌菌悬液离心(5 000 r/min, 15 min ,4℃)。弃去上清液,将沉淀物重新悬浮在生理盐 水中至所需浓度,浓度范围为 108 ~ 109 CFU/mL[17]。1.3.5 过热蒸汽杀菌实验过程 首先设置好过热 蒸汽仪器的温度、流量等参数(温度参数为 170、200、230℃,流量参数为 19、24、29 m³/h ),待 整个系统温度和流量稳定后,关闭阀门,快速将 3根装有 4mL 菌悬液的 18mL 玻璃试管放在物料放 置架上,关闭箱门。重新打开蒸汽阀门,同时记录 杀菌时间(杀菌时间为 40、70、100、150、200、250、300s )。杀菌完成后,将试管立即放入冰水 中冷却,直至微生物计数检测 [18]。 1.3.6 微生物计数 参照方法 1.3.2中菌落总数测 定的方法检测腐败菌在过热蒸汽处理前、后的数 量。过热蒸汽的杀菌效果采用微生物残活率表示,见公式(1)。 过热蒸汽处理前的菌悬液的菌落总数记为 N 0(CFU/mL );过热蒸汽处理后菌悬液的菌落总 数记为 N (CFU/mL )。 1.3.7 杀菌机理的菌悬液制备 按照 1.3.4的方法 配制菌悬液,再按照 1.3.5的方法对菌悬液进行杀 菌,具体的过热蒸汽杀菌条件:过热蒸汽温度 230 ℃、流量 29 m ³/h ,处理时间分别为:0、40、70、100、150、200、250 s 。 1.3.8 菌悬液电导率测量 对按照 1.3.7的方法所 制备的菌悬液,使用电导率仪测定其电导率值。 1.3.9 菌悬液紫外吸收测量 对按照 1.3.7的方法 所制备的菌悬液进行离心(5000r/min,15min ,4 ℃),取上清液,在波长 260nm 处测定其吸光 值。 1.3.10 细菌微观形态的观察 借助扫描电子显微 镜对小龙虾产品的优势腐败菌菌株细胞形态进行 观察。对按照 1.3.7的方法所制备的菌悬液进行离 心(5 000 r/min, 15 min ,4 ℃,弃去上清液,下同),再加入 0.1 mol/L pH =7.0的 PBS 缓冲液,离心。加 入适量的体积分数 2.5%的戊二醛溶液固定过夜后 离心。加入 PBS 缓冲液,离心,重复 3次。分别 加入 30%、50%、70%、80%、90%、100%的乙醇 逐级洗脱,每级洗脱 10 min ,离心,其中 100%的 乙醇洗脱 2次,其余洗脱 1次。再加入无水乙醇与 叔丁醇(体积比=1∶1)洗脱 15 min ,离心。最后 叔丁醇洗脱 15 min ,离心。得到的菌泥再经真空冷 冻干燥后,固定在金属箔片上,喷金 80 s ,电镜观 察。 1.4 杀菌动力学数学模型--Weibull 模型 式中,b 为尺度参数,n 为形状参数,t 为过热 蒸汽处理时间 (s)。当 n = 1,曲线为一条直线,此时 Weiibull 模型为一级动力学方程;当 n >1,曲线 呈凸形;当 n < 1,曲线呈凹形。 1.5 数据分析 每个实验重复 3次,数据用“平均值±标准差”表示,使用 Origin8.5软件对实验数据进行拟合分 析并绘图。 2 结果与分析 2.1 熟制小龙虾贮藏期间菌落总数和挥发性盐基 氮变化 菌落总数直接反映熟制小龙虾产品被细菌污 染的程度和是否符合卫生要求,所以菌落总数在一 定程度上表示小龙虾产品是否具有可食性。挥发性 盐基氮含量表示小龙虾产品由于受到微生物和酶 的作用,本身蛋白质被分解产生氨和胺类等碱性含 氮物质的程度 [19]。图 1是熟制小龙虾在常温下 (25℃)贮藏过程中菌落总数和挥发性盐基氮含 量的变化,由图 1可知,贮藏期间小龙虾菌落总数 一直持续增长,挥发性盐基氮含量先缓慢增加后急 剧上升。孙亚军等 [20]研究南美白对虾虾仁贮藏期间 挥发性盐基氮的变化规律,发现挥发性盐基氮含量 先缓慢增加后快速增加。熟制小龙虾贮藏期间挥发 性盐基氮含量的增长规律与其类似。这是由于贮藏 前期,产品微生物的数量较少,蛋白质被分解的程 度较低,碱性含氮物质的含量较少。随着贮藏时间 延长,微生物数量增加,分解作用变强,从而使挥 发性盐基氮含量增加 [21]。根据 GB 10136-2015《食 品安全国家标准动物性水产制品》,判定熟制小 龙虾达到腐败终点的菌落总数限量为 5×104CFU/g ,挥发性盐基氮限定值为 30 mg/100 g 。贮藏 2 d ,小龙虾的菌落总数为 4.47×105 CFU/g ,已超过 限定值,此时挥发性盐基氮含量为 19.45 mg/100 g 还未超标,挥发性盐基氮指标滞后于菌落总数指 标。综合来看,贮藏 2 d 的熟制小龙虾已腐败变质,到达货架期的终点。 图 1 熟制小龙虾菌落总数和挥发性盐基氮含量 随贮藏时间的变化 Fig. 1 Change in total bacterial count and TVB-N value of cooked crayfish during storage 2.2 小龙虾优势腐败菌的分离与鉴定 根据 1.3.3,从稀释度为 10-3的平板上共挑取 56个菌落,然后分别划线分离纯化。根据图 2可知,小龙虾优势腐败菌构建的系统发育树可判定 A1号 菌为短小芽孢杆菌(Bacillus pumilus );B2号菌为 解淀粉芽孢杆菌(Bacillus amyloliquefaciens );C12号菌为蜡样芽孢杆菌(Bacilluscereus );D4号菌 为赖氨酸芽孢杆菌(Lysinibacillus macroides )。它 19.64%、14.29%、42.86%和 23.21%。 们均为革兰氏阳性菌,并分别占挑取分离总菌落的 图 2 4种优势腐败菌系统发育树 Fig. 2 Phylogenetic tree of four dominant spoilage bacteria 从小龙虾产品中分离出的蜡样芽孢杆菌和赖 氨酸芽孢杆菌与相关文献报道一致 [22]。而解淀粉芽 孢杆菌和短小芽孢杆菌是首次在腐败小龙虾产品 中被发现。在真空包装中,革兰氏阳性菌的数量远 远大于革兰氏阴性菌 [23]。蜡样芽孢杆菌、解淀粉枯 草芽孢杆菌和短小芽孢杆菌均被认为是熟制食品 的优势腐败菌,均在小麦面包 [24]、鱼糜 [25]和烤鸡 [26]等中被发现。因煮制的杀菌温度较低,同时解淀粉 枯草芽孢杆菌和短小芽孢杆菌具有良好耐热性,所 以他们能够在煮制过程中存活并在贮藏期间生长 繁殖。因此解淀粉枯草芽孢杆菌和短小芽孢杆菌能 够在经煮制后的小龙虾产品中大量存在并成为小 龙虾的优势腐败菌。 2.3 过热蒸汽温度和杀菌时间对小龙虾优势腐败 菌的杀菌动力学 研究过热蒸汽流量为 24 m³/h 、不同过热蒸汽温 度和杀菌时间对 4种优势腐败菌的灭活效果,其结 果如图 3所示。在整个杀菌过程,随着杀菌时间延 长,微生物残活率持续下降,直到完全杀灭。但整 个过程,微生物残活率下降的速率是不同的。在杀 菌初始阶段,4种优势腐败菌的残活率均迅速下降,并呈现类似的趋势。然后随着过热蒸汽处理时间的 延长,4种优势腐败菌的残活率变化缓慢,趋于平 缓。此外,过热蒸汽对 4种小龙虾优势腐败菌的杀 菌动力学所拟合的生存曲线均为“拖尾”曲线。这 是由于在过热蒸汽处理初期,过热蒸汽遇见冷试管 时,蒸汽在试管表面冷凝,释放出大量的热量并传 递给菌悬液,使其温度快速升高,从而快速的杀死 菌悬液中存活的细菌。随着过热蒸汽的处理,菌悬 液中存在的热敏性细菌个体被杀灭,留下耐热性较 强的细菌个体,导致残活率下降速率变慢 [27]。上述 趋势与过热蒸汽杀灭嗜热脂肪芽孢杆菌孢子 (Geobacillusstearothermophilus spores )的过程类 似 [28]。这种趋势不仅与杀菌工艺有关,还与细菌细 胞壁有关。4种小龙虾优势腐败菌均为革兰氏阳性 菌。革兰氏阳性菌拥有多层肽聚糖和磷壁酸组成的 坚固和厚实的细胞壁,磷壁酸有助于加强细胞壁的 机械强度,从而提高细菌热耐性 [29]。 杀菌效果也受过热蒸汽温度的影响。过热蒸汽 对 4种小龙虾优势腐败菌的灭活效果与过热蒸汽温 度呈正相关。在相同杀菌时间,过热蒸汽温度为 230℃的杀菌效果要优于 170℃。较高过热蒸汽温 度使蒸汽在冷凝时释放大量的热和较强的气体辐 射,因此使菌悬液的温度上升得更快,具有更高的 微生物杀菌效率 [30]。同时,不同腐败菌对不同过热 蒸汽温度有不同的反应。蒸汽温度 170℃,杀菌时 间为 250 s 时,解淀粉芽孢杆菌的残活率为-6.88,而在相同条件下,其他菌株对应的残活率均<-8。在相同温度条件下,其残活率不同,可能与菌株本 身的耐热性有关。在 4种小龙虾优势腐败菌中,B2菌株的耐热性最强。BOUKHRIS I 等发现解淀粉样 芽孢杆菌具有很强耐热性 [31]。总之,过热蒸汽对 4种小龙虾优势腐败菌有较好的杀菌效果。过热蒸汽 处理(200 ℃,250 s ;230℃,200 s )后,其残活 率均<-8,杀菌效果均能达到近 100%的致死率。 2.4 过热蒸汽流量对小龙虾腐败菌杀菌效果的影响 过热蒸汽温度为 200℃,不同过热蒸汽流量的 杀菌过程中,蒸汽流量对 4种优势腐败菌杀菌效果 的变化趋势与蒸汽温度相似。其结果如图 4所示,在杀菌初期,残活率迅速减小。随处理时间的延长,腐败菌的残活率变化逐渐变缓慢。拟合不同蒸汽流 量对小龙虾优势腐败菌的动力学曲线都呈带有 “拖 尾 ”的曲线。通过过热蒸汽(19 m³/h ,300 s ;24 m³/h ,250 s ;29 m³/h ,200 s )处理后,小龙虾优势腐败菌 能被完全杀灭,其杀菌效果均能达到近 100%的致 死率。 A1(短小芽孢杆菌);B2(解淀粉芽孢杆菌);C12(蜡样芽孢杆菌);D4(赖氨酸芽孢杆菌) A1 (Bacillus pumilus); B2 (Bacillus amyloliquefaciens); C12 (Bacillus cereus); D4 (Lysinibacillus macroides).图 3 在 24 m³/h 蒸汽流量下杀菌处理小龙虾产品优势腐败菌的存活曲线 Fig. 3 Survival curves of dominant spoilage bacteria from crayfish product after different 24 m³/h steam flow rate 19m3/h 实测值 时间 /s 时间 /s A1(短小芽孢杆菌);B2(解淀粉芽孢杆菌);C12(蜡样芽孢杆菌);D4(赖氨酸芽孢杆菌) A1 (Bacillus pumilus); B2 (Bacillus amyloliquefaciens); C12 (Bacillus cereus); D4 (Lysinibacillus macroides) 图 4 在 200℃蒸汽温度下杀菌处理小龙虾产品优势腐败菌的存活曲线 Fig. 4 Survival curves of dominant spoilage bacteria from crayfish product after different superheated steam flow rates, treatment time and 200℃steam temperature 2.5 Weibull 模型参数及拟合评价 运用 Weibull 模型拟合不同过热蒸汽温度、流 量和杀菌时间因素下杀灭 4株小龙虾优势腐败菌后 残活率的曲线。由表 1可知,Weibull 模型的 R 2≥0.935,表明 Weibull 模型对过热蒸汽灭活小龙虾产 品中优势腐败菌的所有动力学都具有较高拟合度。 Weibul 模型因只有 2个参数 b 、n ,具有一定的简洁 性、灵活实用性,可避免过度参数化 [32]。尺度参数 b 反映过热蒸汽参数与杀菌效果间相关性,形状参 数 n 反映曲线的形状。由表 1可知,参数 b 随着过 热蒸汽温度、流量的增大而增大,说明过热蒸汽参 数与杀菌效果呈正相关,所以提高过热蒸汽温度和 增大流量,更容易到达灭活腐败菌的效果;参数 n 的值均小于 1(表 1),表明拟合曲线的形状为先 迅速下降,后趋于平缓,呈凹形。因此,延长杀菌 时间并不能够提高整个杀菌过程的杀菌效率 [33]。为 避免拖尾现象发生、提高过热蒸汽杀菌效果和保障 产品品质,过热蒸汽对小龙虾腐败菌的杀菌过程应 提高过热蒸汽温度、增大流量、缩短杀菌时间。 表 1 Weibull 模型参数和 R 2 Table 1 Weibull model parameters and R 2 菌株编号 温度/℃ b n R2 菌株编号 流量/(m³/h) b n R2 A1 170 0.75 0.44 0.973 19 0.80 0.43 0.982 200 0.99 0.39 0.976 A1 24 1.08 0.37 0.969 230 1.41 0.33 0.971 29 1.21 0.35 0.967 170 0.92 0.36 0.991 19 0.93 0.39 0.968 B2 200 0.89 0.39 0.985 B2 24 1.18 0.35 0.983 230 1.20 0.34 0.980 29 1.42 0.32 0.965 170 0.12 0.75 0.979 19 0.74 0.43 0.935 C12 200 0.40 0.55 0.966 C12 24 0.75 0.44 0.960 230 1.10 0.36 0.978 29 0.76 0.44 0.952 170 1.11 0.38 0.972 19 0.78 0.43 0.963 D4 200 1.22 0.36 0.964 D4 24 1.45 0.32 0.957 230 1.60 0.31 0.971 29 2.22 0.24 0.953 为衡量实际值与预测值的接近程度,以过热蒸 汽温度、流量所有的实测值分别为横坐标,模型预 测值为纵坐标作图,进行线性拟合,其结果如图 5所示。实测值与预测值越接近,其关系曲线的斜率 a 、决定系数 R 2均接近于 1,截距 b 接近于 0。由图 5可知,过热蒸汽温度和流量的线性回归方程分别 为 y = 0.984 11x -0.090 6(R 2 =0.984 98)、y = 0.978 6x -0.127 73(R 2=0.979 31)。斜率 a 、截距 b 分别为 a =0.984 11、0.978 6;b =0.090 6、0.127 73。斜率 a 、R 2均接近 1,截距 b 接近于 0,说明 Weibull 模型得 到的预测值与实测值接近程度很高。 A:温 度 B:流量 图 5 过热蒸汽温度和流量对小龙虾腐败菌杀菌效果的实测值和预测值的相关性 Fig. 5 Correlation between measured value and predicted value for inactivation effects of spoilage bacteria in crayfish by steam temperature and flow rate 2.6 杀菌机理初探 2.6.1 电导率及紫外吸收 细胞膜是维持细胞完 整和细胞正常物质运输、能量代谢的重要结构,当 细菌的细胞膜受到破坏时,细胞膜失去控制物质运 输的功能,细胞内电解质大量外流,导致菌悬液的 电导率升高。同时膜内小分子物质和核酸等大分子 物质也随之流出,这些核酸物质在波长 260nm 处 有吸收 [34]。因此通过测量菌悬液的电导率和 260 nm 吸光度的变化情况,了解菌体在过热蒸汽处理后细 胞膜的受损情况。从图 6-a 可知,0 ~ 40 s ,菌悬液 的电导率数值上升最快,40 ~ 250 s ,菌悬液的电导 率上升的速率比前 40 s 缓慢。如图 6-b 所示,菌悬 液在 260nm 的吸光度随着过热蒸汽处理时间延长 呈不断上升趋势,先快速升高后平缓增加。电导率 与吸光度的变化表明在过热蒸汽处理 40 s 后菌悬液 中已有大量菌体的细胞膜受到破坏 [35]。电导率和紫 外吸收的变化规律与小龙虾产品优势腐败菌的存 活曲线(图 3、图 4)具有一致性。在杀菌初始阶 段,大量优势腐败菌被杀灭,随着过热蒸汽处理时 间延长,过热蒸汽的杀菌速率变缓慢,最后趋于稳 定。 A1(短小芽孢杆菌);B2(解淀粉芽孢杆菌);C12(蜡样芽孢杆菌); D4(赖氨酸芽孢杆菌) A1 (Bacillus pumilus ); B2 (Bacillus amyloliquefaciens); C12 (Bacillus cereus ); D4 (Lysinibacillus macroides ). 图 6 小龙虾产品优势腐败菌电导率和紫外吸收随过热蒸 汽处理时间的变化 Fig. 6 The changes of conductivity and UV absorption of the dominant spoilage bacteria from crayfish product treated with superheated steam treatment time 2.6.2 细菌微观形态的观察 图 7表示小龙虾优势 腐败菌蜡样芽孢杆菌的微观形态随过热蒸汽处理 时间的变化。从图 7-A 观察可知,未经过热蒸汽处 理的细胞具有规则的长杆状,形态丰满、完整、未 变形,细胞表面光滑,无破裂,无内容物外泄现象。过热蒸汽处理 40s (图 7-B )后,菌体保持较完整 的杆状,但细胞表面变粗糙,出现不规则的褶皱, 表现为凹凸不平,同时存在内容物外泄现象,此时 菌体形态已遭到破坏,细胞膜或细胞壁已破裂,导 致菌悬液的电导率和紫外吸收数值的增加。过热蒸 汽处理 150 s 和 250 s 后(图 7-C 和图 7-D ),菌体 形态扭曲变形,菌体四周不完整,破损和断裂严重,甚至有些菌体破裂成细胞碎片。结果表明,随过热 蒸汽处理时间延长,改变了菌株外部形态,并对细 菌的细胞壁膜造成不可逆的损伤,以及胞内物质外 流,从而导致菌体死亡。 图 7 过热蒸汽分别处理 0、40、150、250 s 蜡样芽孢杆菌 的扫描电镜图 Fig. 7 Scanning electron microscopy of Bacillus cereus at different inactivation time with superheated steam 3 结论 采用稀释涂布平板法,从腐败的小龙虾产品中 分离出优势腐败菌,经重复划线纯化得到纯菌落。根据优势腐败菌的 16S rDNA 基因序列将其鉴定为 短小芽孢杆菌(Bacillus pumilus )、解淀粉芽孢杆 菌(Bacillusamyloliquefaciens )、蜡样芽孢杆菌 ( Bacillus cereus )和赖氨酸芽孢杆菌 (Lysinibacillus macroides )。Weibull 模型能较好拟 合过热蒸汽温度、流量和杀菌时间对熟制小龙虾优 势腐败菌的灭活动力学,其 R 2≥0.935。经过热蒸 汽处理后,4种小龙虾产品的优势腐败菌均能被完 全杀灭。提高过热蒸汽温度和流量可以使过热蒸汽 对熟制小龙虾腐败菌的整体杀菌效果得到提升。同 时通过对过热蒸汽处理后菌悬液电导率和吸光度 测量,以及对菌体微观结构的观察,表明过热蒸汽 杀菌使菌体细胞膜遭到严重变化,使细菌形态发生 变化,造成胞内物质外流,致使腐败菌死亡。本研 究结果证实过热蒸汽是在小龙虾产品加工杀菌过 程中一种有前途的杀菌技术。 参考文献 [1] 李锐 ,江祖彬 ,童光森 ,等 .即食麻辣小龙虾加工工艺 研究 [J].食品研究与开发 , 2019, 40(5):138-143. [2] 肖放 ,刘忠松 ,郭云峰 ,等 .中国小龙虾产业发展报告 (2017)[J].中国水产 , 2017(7): 8-17. [3] 江杨阳 .小龙虾低温贮藏品质变化规律及其腐败菌鉴 定 [D].杭州 :浙江大学 , 2019. [4] 葛孟甜 ,李正荣 ,赖年悦 ,等 .两种杀菌方式对即食小 龙虾理化性质及挥发性风味物质的影响 [J].渔业现代化 ,2018, 45(3): 66-74. [5] KOWN S A, SONG W J, KANG D H. Comparison of the effect of saturated and superheated steam on the inactivationof Escherichiacoli O157:H7,Salmonella Typhimurium and Listeria monocytogenes on cantaloupe and watermelon surfaces[J]. Food Microbiology, 2018, 72:157-165. [6] 王瑞睿 .过热蒸汽加工对猪肉品质影响研究 [D].北京 :中国农业科学院 , 2019. [7] GA-HEE B, DONG-HYUN K. Inactivation of Escherichia coli O157:H7, Salmonella typhimurium , and Listeria monocytogenes on cherry tomatoes and oranges by superheated steam[J]. Food Research International, 2018,112: 38-47. [8] HEADDS,CENKOWSKIS,HOLLEYR.Effectsof superheated steam on Geobacillus stearothermophilus spore viability[J]. Journal of Applied Microbiology, 2008, 104(4):1213-1220. [9] BAN G H , KANG D H . Effectiveness of superheated steam for inactivation of Escherichia col i O157:H7, Salmonella Typhimurium,Salmonella Enteritidisphage type30,and Listeria monocytogenesonalmondsand pistachios[J]. International Journal of Food Microbiology,2016, 220: 19-25. [10] ARTIGUEZ M L. Inactivation of spores and vegetative cells of Bacillus subtilis and Geobacillus stearothermophilus by pulsed light[J]. Innovative Food Science & Emerging Technologies, 2015, 28: 52-58. [11] CARMEN O, QUILES A , BENEDITO J. Inactivation kinetics and cell morphology of E. coli and S. cerevisiae treatedwithultrasound-assistedsupercriticalCO2[J]. Food Research International, 2014, 62: 955-964. [12] ÉRICASS,JORGEAWG,ANTONIOM,etal. Inactivation kinetics of Escherichia coli O157:H7 and Listeria monocytogenes in apple juice by microwave andconventionalthermalprocessing[J].Innovative FoodScience&EmergingTechnologies,2018,45:84-91. [13] BAN C, LEE D H, JO Y, et al. Use of superheated steam to inactivate Salmonella enterica serovars Typhimurium and Enteritidis contamination on black peppercorns, pecans, and almonds[J].Journal ofFoodEngineering,2018, 222: 284-291. [14] 张泽伟 ,吉宏武 ,段伟文 ,等 .两种灭菌方式对熟制 小龙虾冷藏期间品质的影响 [J].广东海洋大学学报 ,2019, 39(06):93-100. [15] 童依婷 ,晏冬华 ,彭文煊 ,等 .朱蕉叶枯病菌的鉴定 及生物学特性 [J].广东海洋大学学报 ,2016,36(4):89-95. [16] 叶章颖 ,祁凡雨 ,裴洛伟 .微酸性电解水对虾仁的杀 菌效果及其动力学 [J].农业工程学报 ,2014,30(3):223-230. [17] 雷玉洁 ,周良付 ,李宇坤 ,等 .大肠杆菌射频加热杀 菌效果及动力学模型拟合 [J].中国食品学报 ,2018,18(4):132-138. [18] LVR,CHANTAPAKULT,ZOUM,etal.Thermal inactivation kinetics of Bacillus cereus in Chinese rice wine and in simulated media based on wine components[J]. Food Control, 2018, 89: 308-313. [19] 吴湛霞 ,潘江球 ,杨子明 ,等 .混合分子质量壳聚糖 结合臭氧杀菌处理对罗非鱼片的冷藏保鲜作用 [J].广 东海洋大学学报 , 2016, 36(3): 71-75. [20] 孙亚军 ,雷晓凌 ,钟敏 .冷鲜虾仁中一株优势腐败菌 的腐败品质动力学研究 [J].食品与机械 ,2014(1):155-158. [21] 丁捷 ,胡欣洁 ,卢雪松 ,等 .糊辣牛肉在不同温度条 件下的贮藏特性及其货架期预测 [J].现代食品科技 ,2017, 33(03): 122-132;104. [22] 周涛 ,宋方霞 ,黄村惠 .克氏原螯虾调理产品中主要 腐败菌的分离鉴定 [J].南京师大学报 (自然科学版 ),2017, 40(4): 103-108;115. [23] ZHANGYM,LIQ,LIDP,etal.Changesinthe microbial communities of air-packaged and vacuum-packaged common carp (Cyprinus carpio ) stored at 4 °C[J]. Food Microbiology, 2015, 52:197-204. [24] VALERIO F, BELLIS P D, BIASE M D, et al. Diversity of spore-forming bacteria and identification of Bacillus amyloliquefaciens asaspeciesfrequentlyassociated with the ropy spoilage of bread[J]. International Journal of Food Microbiology, 2012, 156(3): 278-285. [25] LYVER A , SMITH J P , AUSTIN J, et al. Competitive inhibition of Clostridium Botulinum type E by Bacillus speciesin avalue-addedseafoodproductpackaged under a modified atmosphere[J]. Food Research International, 1998, 31(4):311-319. [26] 徐世明 ,赵瑞连 ,宋维娟 .胀袋烤鸡中腐败菌的分离 与鉴定 [J].食品科技 , 2011, 36(2): 13-15. [27] BUZRUL S, ALPAS H, BOZOGLU F. Use of Weibull frequency distribution model to describe the inactivation of Alicyclobacillusacidoterrestris by high pressure at different temperatures[J]. Food Research International,2005, 38(2): 151-157. [28] CENKOWSKI S, PRONYK C, ZMIDZINSKA D, et al. Decontamination of food products with superheated steam[J].JournalofFoodEngineering,2007,83(1):68-75. [29] ZHANG Y X, WEI J P, YUAN Y H, et al. Bactericidal effect of cold plasma on microbiota of commercial fish balls [J]. Innovative Food Science & Emerging Technologies, 2019, 52: 394-405. [30] HU Y M, NIE W , HU X Z, et al. Microbial decontamination of wheat grain with superheated steam[J]. Food Control, 2016, 62: 264-269. [31] BOUKHS I, FARHAT-KHEMAKHEM A, BLIBECH M, et al. Characterization of an extremely salt-tolerant and thermostablephytasefrom Bacillusamyloliquefaciens US573[J]. International Journal of Biological Macromolecules, 2015, 80: 581-587. [32] BARANYIJ,ROSST,ROBERTSTA.Effectsof parameterization on the performance of empirical models used in`predictive microbiology' [J]. Food Microbiology, 1996, 13(1): 83-91. [33] 李珊 ,陈芹芹 ,李淑燕 .超高压对鲜榨苹果汁的杀菌 效果及动力学分析 [J].食品科学 , 2011, 32(7): 43-46. [34] 刘书成 ,郭明慧 ,刘媛 ,等 .高密度 CO2杀菌和钝酶 及其在食品加工中应用的研究进展 [J].广东海洋大学 学报 , 2016, 36(4): 101-116. [35] 刘蔚 ,周涛 .ε-聚赖氨酸抑菌机理研究 [J].食品科学 ,2009, 30(9): 15-20. (责任编辑 :刘岭)
确定






还剩8页未读,是否继续阅读?

产品配置单





中国格哈特为您提供《过热蒸汽杀菌熟制小龙虾中挥发性盐基氮的测定》,该方案主要用于熟制动物性水产制品中挥发性盐基氮检测,参考标准--,《过热蒸汽杀菌熟制小龙虾中挥发性盐基氮的测定》用到的仪器有格哈特全自动凯氏定氮仪VAPODEST 450、格哈特快速干燥仪STL56、格哈特强力高重现振荡器LS500/RO500、德国移液器MM、1200ml大体积样品蒸馏管
推荐专场












相关方案
更多
该厂商其他方案
更多