








方案详情
文
在缺少动态信息的情况下,代谢组学数据的解读通常十分复杂,这是因为在相应代谢物丰度未发生改变时,某个通路的代谢流可能已经发生显著变化。稳定同位素示踪(定性代谢流分析)的巨大潜力有助于解决这些状况,以便更深入地了解生物系统。在定性代谢流分析中,将稳定同位素示踪剂(通常含有 13C、15N 或 2H)引入生物系统, 导致下游代谢物的同位素模式(同位素异数体分布)发生改变。该方法尤其适用于易于 引入稳定同位素示踪剂的细胞模型,对产生的标记模式及示踪剂掺入动力学的分析将提供对酶功能、途径依赖、基因表达和蛋白功能变化效应的洞察。 Agilent MassHunter VistaFlux 软件旨在设计一套完整的工作流程,帮助科学家进行定性代谢流分析,助于高分辨率准确质量数 TOF 和 Q-TOF LC/MS 数据的处理和可视化。软件提供的工具可进行特征提取、同位素掺入分析、同位素异数体丰度分析、天然丰度校正、统计学分析以及数据在通路中的可视化。
方案详情

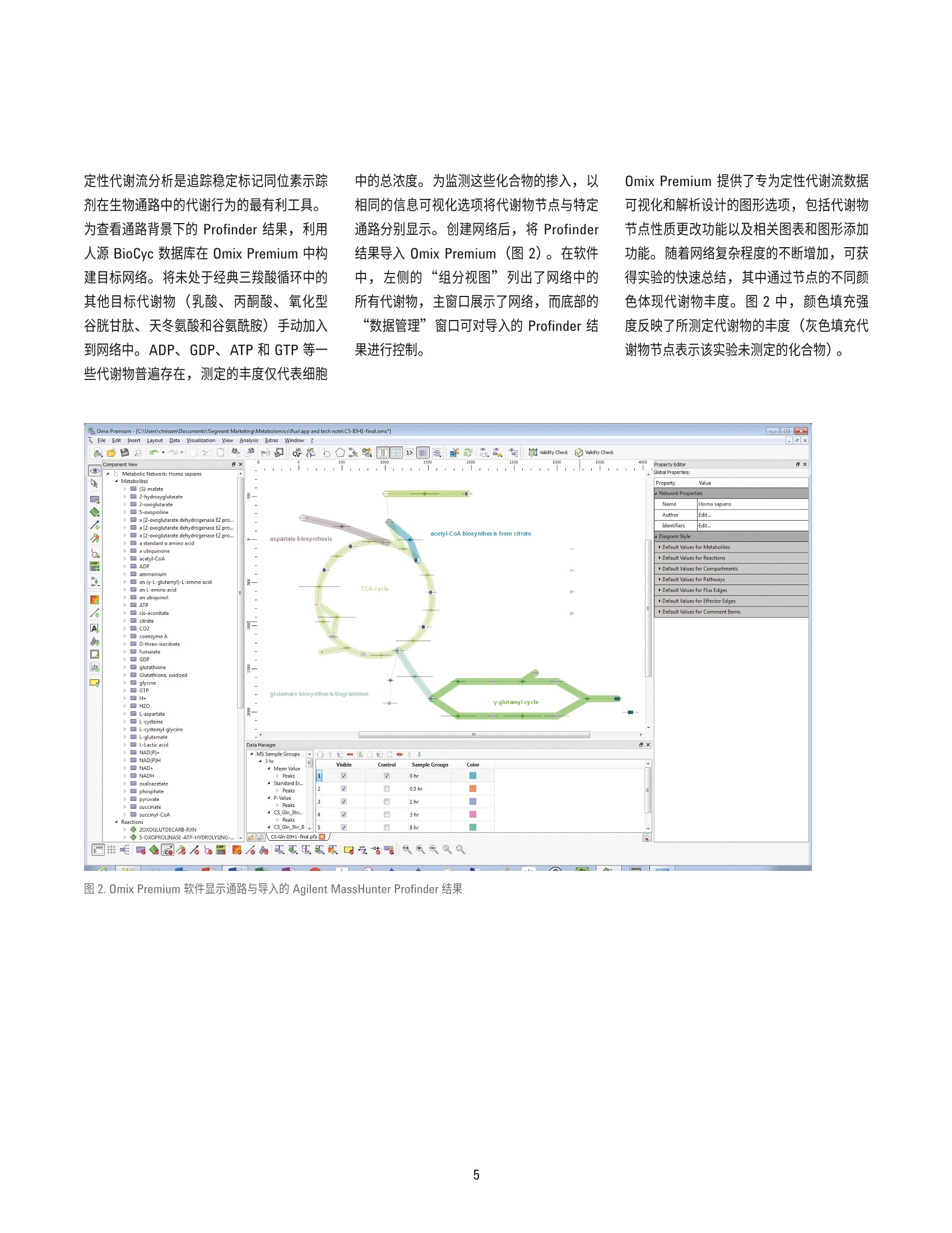
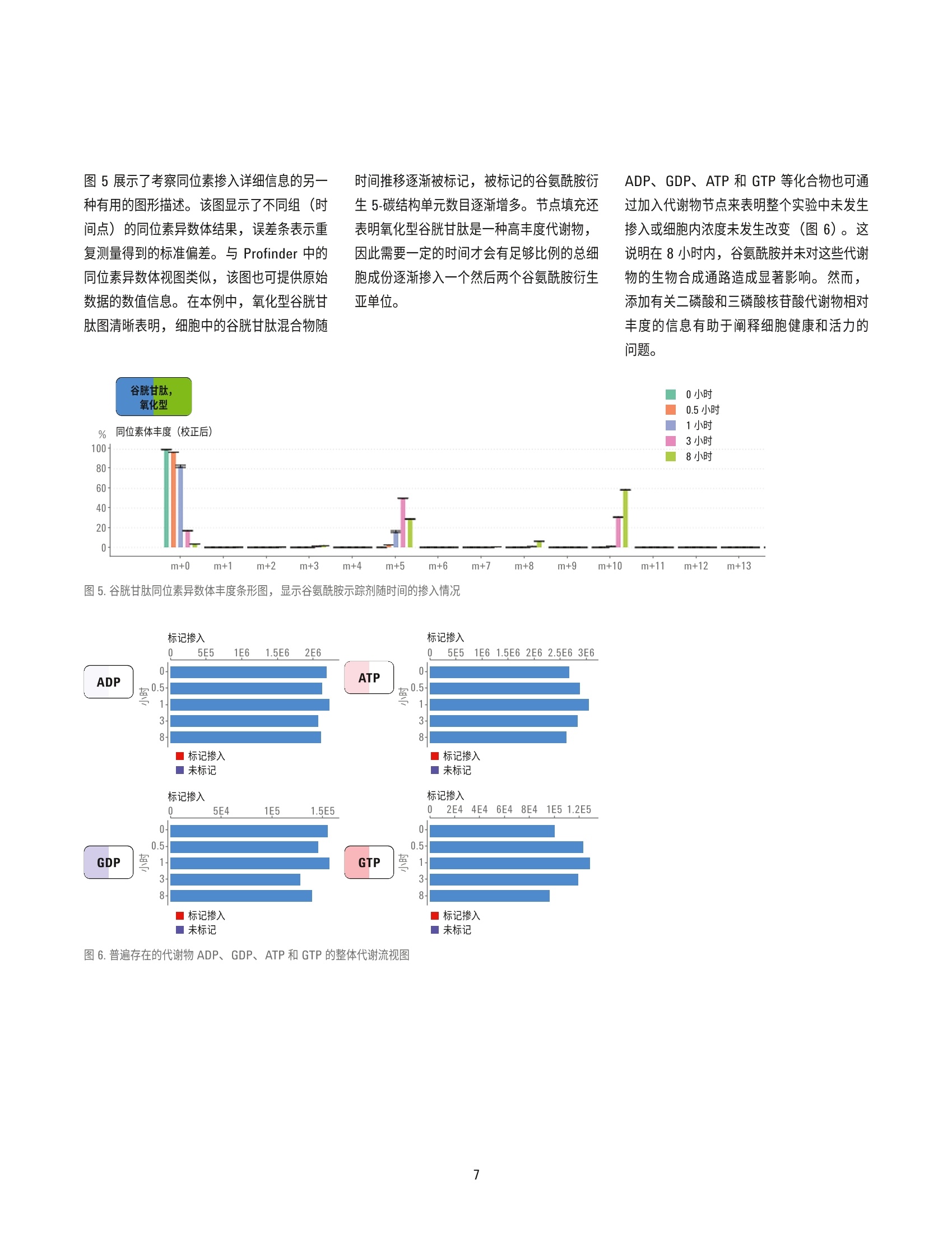
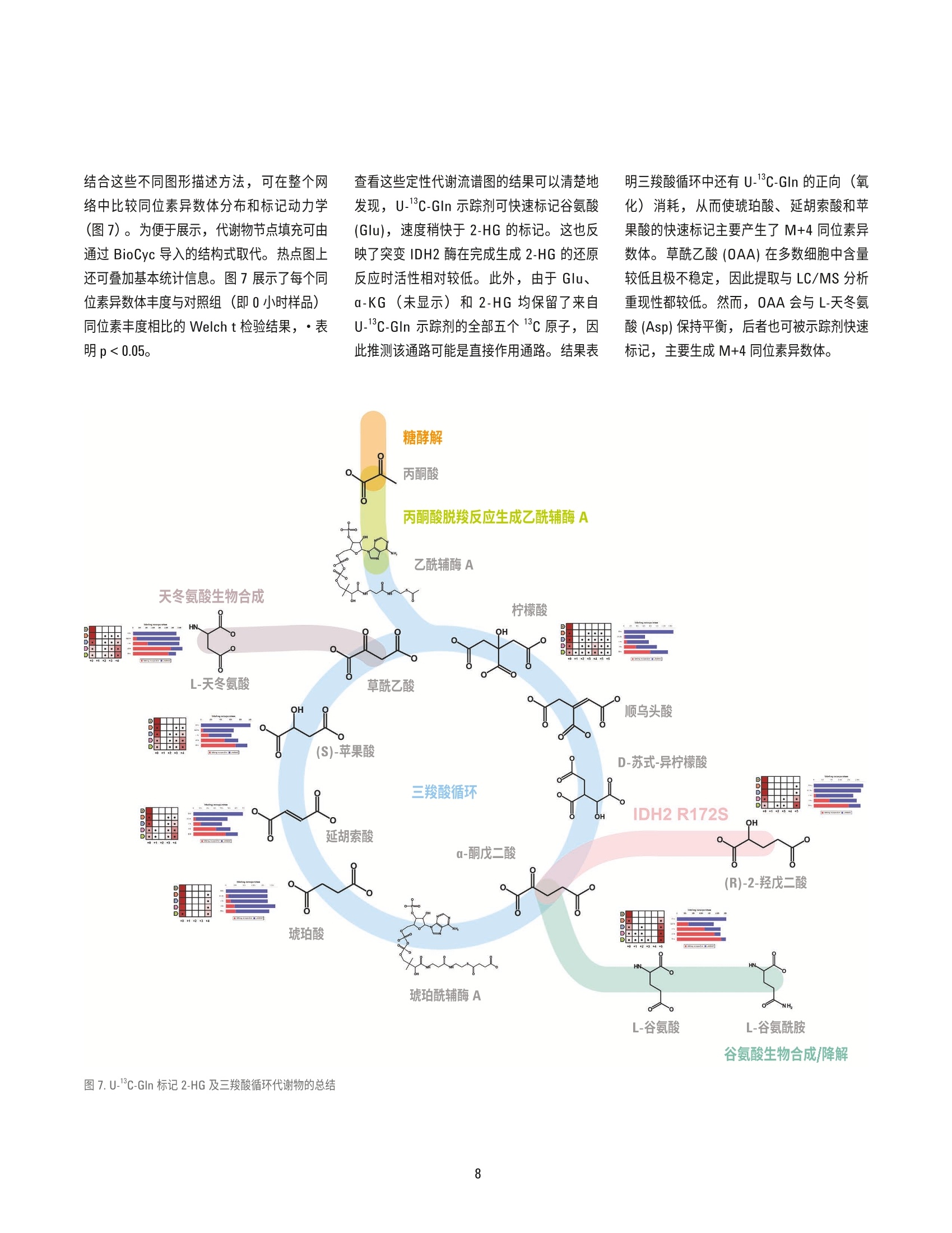
( 作者 ) Hui Liu、Weige Qin、Hardik Shah、 Bhavapriya Vaitheesvaran、 ( Vladimir Yong 和 Justin R. Cross ) Donald B. 和 Catherine C. Marron 癌症代谢中心, 纪念斯隆-凯特琳癌症中心 Lucia Salamanca-Cardona 和 Kayvan R. Keshari 放射学与分子药理学项目部, 纪念斯隆-凯特琳癌症中心 使用 Agilent VistaFlux 对软骨肉瘤细胞系进行13C-谷氨酰胺定性代谢流分析 应用简报 代谢组学 摘要 在缺少动态信息的情况下,代谢组学数据的解读通常十分复杂,这是因为在相应代谢物丰度未发生改变时,某个通路的代谢流可能已经发生显著变化。稳定同位素示踪(定性代谢流分析)的巨大潜力有助于解决这些状况,以便更深入地了解生物系统。在定性代谢流分析中,将稳定同位素示踪剂(通常含有C、15N或H)引入生物系统,导致下游代谢物的同位素模式(同位素异数体分布)发生改变。该方法尤其适用于易于引入稳定同位素示踪剂的细胞模型,对产生的标记模式及示踪剂掺入动力学的分析将提供对酶功能、途径依赖、基因表达和蛋白功能变化效应的洞察。 Agilent MassHunter VistaFlux 软件旨在设计一套完整的工作流程,帮助科学家进行定性代谢流分析,助于高分辨率准确质量数 TOF 和 Q-TOF LC/MS 数据的处理和可视化。软件提供的工具可进行特征提取、同位素掺入分析、同位素异数体丰度分析、天然丰度校正、统计学分析以及数据在通路中的可视化。 本应用简报展示了使用13C-谷氨酰胺(U-3C-GIn)作为示踪剂,对软骨肉瘤细胞系进行定性代谢流分析的研究示例。采用安捷伦 LC/TOF 系统进行数据采集,并使用MassHunter VistaFlux 进行数据分析和解析。研究证明了异柠檬酸脱氢酶2(IDH2)的突变对三羧酸循环代谢产生的影响,特别是突变的 IDH2异构体将致使谷氨酰胺转化生成2-羟基戊二酸(2-HG)。 前言 近期,多种实体瘤和血液肿瘤中均发现了体细胞 IDH 突变, 其中包括胶质瘤、急性髓性白血病和软骨肉瘤。异柠檬酸脱氢酶(IDH1/2) 的突变发生在酶的活性位点上,形成的功能获得性表型导致代谢物2-羟戊二酸(2-HG)的生成能力提高。近期研究表明,IDH突变细胞中 2-HG的高水平累积与清除途径的饱和相关,因此 2-HG 成为了 DNA 脱甲基酶 TET 家族、组氨酸脱甲基酶 Jumonji C 家族、mRNA脱甲基酶 FTO 和ALKBH5 以及调节缺氧诱导因子 (HIF)的脯氨酰羟化酶 (PHD)等各种α-酮戊二酸 (a-KG)依赖型加双氧酶的竞争性抑制剂。这些变化促进了抑制性染色质环境的形成,这种环境可将细胞锁定在未分化状态下。 软骨肉瘤是由软骨细胞沉积软骨引起的一种肉瘤, 具有较高的 IDH突变发生率。软骨肉瘤肿瘤来源细胞系的易得性使其成为了解IDH 突变后果的实用模型系统。其中一项重要研究是使用定性代谢流分析来说明突变酶产生的2-HG 来自 a-KG 和三羧酸循环中的谷氨酰胺。 实验部分 细胞培养和试剂 人软骨肉瘤CS-1细胞系,采用 DMEM培养基,其中含有10%FBS、25 mM 葡萄糖、4mML-谷氨酰胺、100单位/mL 青霉素以及100pg/mL链霉素,置于填充5% CO,的37℃培养箱中。代谢示踪实验中,在标记阶段开始前2天,以300000细胞/孔点样于标准6孔培养板上,每个时间点重复点样三个孔。含有示踪剂的培养基为不含谷氨酰胺的 DMEM, 并向其中加入10%经透析的 FBS 及4mM [U-C]-L-谷氨酰胺 (Isotec 605-166, Sigma)。标记阶段开始时,用 PBS清洗细胞一次,并在收集前的0.5、1、3和8小时分别将其转移至含有示踪剂的培养基中。由于培养基的改变也会对代谢组产生干扰,因此对照组(未标记)细胞的收集为在将培养基改变为含10%经透析的 FBS 及4mML-谷氨酰胺(未进行同位素富集)的 DMEM 8小时后进行,此时对照细胞与经过 U-13C-GIn 8 小时处理后的培养基完全匹配。 代谢物提取 轻轻吸出培养基,将细胞置于1 mL/孔 80%(体积比)甲醇水溶液中进行提取。孔板在-80°℃下孵育1小时以上以促进蛋白质沉淀,然后用细胞刮收集。将提取液转移至微量离心管中,在4°℃下以 21000×g离心15分钟。然后将提取液转移至干净的试管中,并在真空浓缩仪 (Genevac) 中蒸干。 需要注意的是,由于改变细胞培养基及用PBS清洗均会干扰快速交换的代谢组,因此在该定性代谢流实验中,仅在加入含有稳定同位素的培养基之前用 PBS 清洗细胞。收集细胞时,吸出培养基后无需额外清洗即直接加入提取溶剂,以便最大程度保持细胞内代谢物浓度与相关标记模式。虽然如此,使用以上流程时仍需要特别注意培养基中浓度较高代谢物的结果解析,这是因为,与收集细胞前立即用 PBS 清洗的方法相比,这种方法将会使细胞提取物受到更高程度的残留培养基污染。 LC/MS分析 用120 pL水对干燥提取物进行重悬,置于冰上孵育20分钟,每5分钟涡旋混合一次。然后在4℃下以21000×g离心15分钟,以去除残留颗粒物。将100 pL提取物转移至体积更小的自动进样器样品瓶中,每个样品中取 10 pL 组成混合 QC样品,分析运行前对该 QC样品进样六次,然后定期在整个分析中间隔进样,以确保色谱稳定性。 ( 由安捷伦液质联用系统采集数据,其中包括: ) ( Agilent 6230 飞行时间质谱仪 ) ( Agilent 1290 Infinity 二元 UHPLC 泵 ) ( Agilent 1290 Infinity 柱温箱 ) ( Agilent 1290 Infinity HTS 自动进样器 ) 使用配备 100:1分流器的 Agilent 1260Inifinity 等度泵引入参比质量溶液。使用双电喷雾离子源连接液相色谱与质谱,在负离子模式下采用 4 GHz 高分辨率,,以每秒1.5张谱图的速率采集数据。表1展示了分析方法设置。 在本实验中,采用目标代谢物分析方法检测。由于三羧酸循环中的有机酸在反相色谱中保留较差,因此采用离子对反相色谱。选择负离子电喷雾离子化模式以获得目标化合物的最佳检测性能。 数据分析 使用 Agilent MassHunter Pathways toPCDL 软件创建了提取自 BioCyc的三羧酸(TCA)循环的安捷伦个人化合物数据库与谱库文件(.cdb)。此外,还在以安捷伦METLIN PCDL 为来源的定制 PCDL 中添加了乳酸、丙酮酸、天冬氨酸、谷氨酸、氧化型谷胱甘肽和2-HG。使用安捷伦 PCDLManager 软件对以上内容进行编辑,以去除分析方法未检出的代谢物,并为每个化合物添加保留时间信息。然后在 AgilentMassHunter Profinder 软件中使用 PCDL文件作为目标列表进行同位素异数体批量提取(表2)。将经过C天然丰度校正的Profinder 批处理结果以 Profinder 存档文件(.PFA)形式导出至 Omix Premium, 文件中包括样品组信息、化合物描述符和同位素异数体丰度。还支持将整个数据集导出为逗号分隔值(.csv) 文件。 参数 值 液相色谱条件 色谱柱 Waters Cortecs C18+ 150 mm×2.1 mm, 2.7 pm 流动相 A)5 mM N,N-二甲基辛胺水溶液, pH 5.5 B)5mMN,N-二甲基辛胺的90%甲醇水溶液, pH 5.5 流速 0-8分钟 0.4 mL/min, 8.1-10分钟 0.6 mL/min 梯度 0 min 寸 B 为 10% 8.0 min 时B为100% 停止时间 10 min 后运行时间 5 min 柱温 30°C 进样量 15 pL 质谱条件 电离模式 双 ESI离子源 电离极性 负 干燥气温度 250°℃ 干燥气 13L/min 雾化器压力 45 psi 毛细管电压 3500V 八极杆1RF电压 600 V 采集范围 m/z 50-1700 表2.用于同位素异数体提取的目标代谢物列表 化合物 分子式 CAS RT (min) L-谷氨酸 CgH,NO, 56-86-0 2.26 L-天冬氨酸 CH,NO 56-84-8 2.29 L-乳酸 C3Hg0: 79-33-4 2.93 谷胱甘肽,氧化型 C20H32N012S2 27025-41-8 4.84 琥珀酸 CAH,04 110-15-6 5.05 2-羟戊二酸 CgHg0s 2889-31-8 5.11 富马酸 CH 04 110-17-8 5.13 L-苹果酸 CAH,05 97-67-6 5.13 酮戊二酸(a-KG) C,H,0, 328-50-7 5.24 二磷酸鸟苷(GDP) C10H15N,01P2 146-91-8 5.83 二磷酸腺苷 (ADP) C10H15N,010P2 58-64-0 5.93 乌头酸 CgH 06 499-12-7 6.1 苏式-异柠檬酸(未与柠檬酸分离) CgHg07 6061-97-8 6.1 柠檬酸(未与苏式-异柠檬酸分离) C,Hg0 77-92-9 6.1 三磷酸鸟苷 (GTP) C10H16Ng014P3 86-01-1 6.57 三磷酸腺苷 (ATP) C10H16N,013P3 987-65-5 6.63 在 Profinder 中完成同位素异数体提取后,对数据进行查看,必要时进行手动调整;只有谷氨酸和 a-KG 需要通过手动调整以获得一致的结果。 Profinder 中的同位素异数体视图为稳定标记的掺入以及组内平行样品的重现性提供了有用的洞察。可以按照原始值或百分比值显示,可以按照经过或未经过天然同位素丰度额外校正显示。图1为2-HG 的同位素异数体结果。所展示的结果为以时间点分组的天然丰度校正值。在本例中,不同时间点的生物学平行分析结果具有良好的一致性,但需要时可在此阶段将异常值样品排除。数据的初步检验结果表明,2-HG M+0 随时间减少,而 2-HG M+5 相应增多,说明 2-HG的所有五个碳均来源于U-13C-GIn 示踪剂,而8小时后多数2-HG代谢物呈标记状态,表明该代谢物来源于谷氨酰胺。 图 1. Agilent MassHunter Profinder 的 2-HG 同位素异数体视图 定性代谢流分析是追踪稳定标记同位素示踪剂在生物通路中的代谢行为的最有利工具。为查看通路背景下的 Profinder 结果,利用人源 BioCyc 数据库在 0mix Premium 中构建目标网络。将未处于经典三羧酸循环中的其他目标代谢物(乳酸、i丙酮酸、氧化型谷胱甘肽、天冬氨酸和谷氨酰胺)手动加入到网络中。ADP、GDP、ATP 和 GTP 等一些代谢物普遍存在,测定的丰度仅代表细胞 中的总浓度。为监测这些化合物的掺入,以相同的信息可视化选项将代谢物节点与特定通路分别显示。创建网络后, 将 Profinder结果导入 Omix Premium (图2)。在软件中,左侧的“组分视图”列出了网络中的所有代谢物,主窗口展示了网络,而底部的“数据管理”窗口可对导入的 Profinder 结果进行控制。 Omix Premium 提供了专为定性代谢流数据可视化和解析设计的图形选项,包括代谢物节点性质更改功能以及相关图表和图形添加功能。随着网络复杂程度的不断增加, 可获得实验的快速总结,其中通过节点的不同颜色体现代谢物丰度。图2中,颜色填充强度反映了所测定代谢物的丰度(灰色填充代谢物节点表示该实验未测定的化合物)。 代谢物节点的颜色有三个选项,如图3所示的 2-HG。节点填充可表示该代谢物浓度较高(图3A中的颜色强度),或表示发生了稳定标记掺入(图3B)。图 3C 为组合视图,展示上述两类信息。 代谢物节点显示出的信息旨在用于表明与对照组相比,何处节点具有最大的变化倍数。一旦基于整个通路的数据概览审查完成后,便可对稳定同位素掺入进行更详细的考查。图4展示的进一步的信息以热点图和水平条形图显示在每个代谢物节点的旁边。这些图总结了同位素异数体丰度和示踪剂掺入的结果,并扩充了节点处显示的总结信息。在本例中,时间点结果显示在热点图的y轴上,同位素异数体结果显示在x轴上。填充颜色强度代表示踪剂掺入的相对丰度。条形图提供了总的示踪剂掺入到总代谢物(检测到的所有同位素异数体的总和)的丰度比例的另一种示图。这些图形证实了在 Profinder 中手动查看数据时首次观察到的结果,也就是说,用U-1C-GIn孵育3小时和8小时后,2-HG 的M+5同位素异数体中发生了明显的示踪剂标记掺入,证明谷氨酰胺是 IDH2突变的 CS-1细胞中生成 2-HG 的主要底物。 图3.2-HG节点,填充为A) 该代谢物与其他检出代谢物的相对丰度,B)表示为颜色梯度的稳定标记掺入百分比, C)分别填充,左侧为丰度,右侧为掺入百分比 图4. CS-1胞胞系中U-1C-GIn 示踪剂掺入柠檬酸的详细信息 图5展示了考察同位素掺入详细信息的另一种有用的图形描述。该图显示了不同组(时间点)的同位素异数体结果,误差条表示重复测量得到的标准偏差。与 Profinder 中的同位素异数体视图类似,该图也可提供原始数据的数值信息。在本例中,氧化型谷胱甘肽图清晰表明,细胞中的谷胱甘肽混合物随 时间推移逐渐被标记,被标记的谷氨酰胺衍生5-碳结构单元数目逐渐增多。节点填充还表明氧化型谷胱甘肽是一种高丰度代谢物,因此需要一定的时间才会有足够比例的总细胞成份逐渐掺入一个然后两个谷氨酰胺衍生亚单位。 ADP、GDP、ATP 和 GTP 等化合物也可通过加入代谢物节点来表明整个实验中未发生掺入或细胞内浓度未发生改变(图6)。这说明在8小时内,谷氨酰胺并未对这些代谢物的生物合成通路造成显著影响。然而,添加有关二磷酸和三磷酸核苷酸代谢物相对丰度的信息有助于阐释细胞健康和活力的问题。 图5.谷胱甘肽同位素异数体丰度条形图,显示谷氨酰胺示踪剂随时间的掺入情况 图6.普遍存在的代谢物 ADP、GDP、ATP 和 GTP 的整体代谢流视图 结合这些不同图形描述方法,,!可在整个网络中比较同位素异数体分布和标记动力学(图7)。为便于展示,代谢物节点填充可由通过 BioCyc 导入的结构式取代。热点图上还可叠加基本统计信息。图7展示了每个同位素异数体丰度与对照组(即0小时样品)同位素丰度相比的 Welch t 检验结果,.表明p<0.05。 查看这些定性代谢流谱图的结果可以清楚地发现, U-1C-GIn 示踪剂可快速标记谷氨酸(Glu), 速度稍快于 2-HG 的标记。这也反映了突变 IDH2酶在完成生成 2-HG的还原反应时活性相对较低。此外,由于 Glu、a-KG(未显示)和2-HG 均保留了来自U-13C-GIn 示踪剂的全部五个13c原子,因此推测该通路可能是直接作用通路。结果表 明三羧酸循环中还有U-13C-GIn的正向(氧化)消耗,从而使琥珀酸、延胡索酸和苹果酸的快速标记主要产生了M+4 同位素异数体。草酰乙酸 (OAA) 在多数细胞中含量较低且极不稳定,因此提取与 LC/MS分析重现性都较低。然而, OAA 会与L-天冬氨酸(Asp)保持平衡,后者也可被示踪剂快速标记,主要生成 M+4同位素异数体。 随后可在2-HG、琥珀酸、延胡索酸、苹果酸和天冬氨酸中观察到 M+3标记。这来源于M+4 标记的 OAA 以及来自乙酰辅酶A的两个未富集碳,从而产生了柠檬酸M+4。下游代谢物中M+3同位素异数体的产生来源于通过野生型 IDH反应, 以正向(氧化方向)发生的脱羧反应。另外还有证据表明柠檬酸中的 M+5 标记来源于由野生型 IDH 反向作用催化的还原羧化反应。8小时后可观察到更为复杂的标记模式,这是由于: 进一步的三羧酸循环反应 琥珀酸、延胡索酸和柠檬酸对称性导致的标记模式混乱 苹果酸酶的潜在作用 可使用另一种稳定同位素示踪剂或在特定碳位置进行谷氨酰胺标记,以对这些模式进行进一步研究。总之,概要视图能够评价代谢物标记的相对动力学参数,由此可见天冬氨酸和谷氨酸标记在标记8小时后趋于稳定。其他代谢物如需达到代谢稳态,可能需要更 结论 采用Agilent MassHunter VistaFlux 工作流程,在 CS-1软骨肉瘤细胞中以 U-13C-GIn作为代谢示踪剂进行定性代谢流分析。该分析明确证实了 IDH2突变异构体对谷氨酰胺衍生的 a-KG 到 2-HG 的转化活性。8小时后, IDH 突变细胞中大部分 2-HG组均被标记为M+5同位素异数体。IDH2 R172S突变酶的新生变形活性超过了内源性 2-HG脱氢酶,因此 2-HG 的生成量有所增加,从而使 2-HG的累积浓度升高。因此,2-HG成为了 DNA 脱甲基酶 TET 家族和组氨酸脱甲基酶 Jumonji C 家族等多种 a-KG 依赖型加双氧酶的竞争抑制剂,造成了包括 DNA超甲基化在内的表观遗传学变化,促进了未分化染色质环境的产生。分析还展示了 U-1C-GIn示踪剂进入经典三羧酸循环通路中的结果。 与手动数据挖掘挖比, VistaFlux 工作流程为稳定同位素示踪数据处理提供了全面、自动化且快捷的框架,其中包括同位素异数体提取和定性代谢流文件数据的通路可视化。 ( 参考文献 ) ( 1. Losman,J. A.; Kaelin, W. G. Jr. What a difference a hydroxyl makes: mutant IDH,(R)-2-hydroxyglutarate, and cancer. Genes Dev. 2013,2 7 (8), 8 36-52 ) ( 2. S usa, M.; et al. Alendronate inhibits growth of high-grade chondrosarcoma cells. Anticancer Res. 2009,29, 1879-1888 ) 查找当地的安捷伦客户中心: www.agilent.com/chem/contactus-cn 免费专线: 800-820-3278,400-820-3278(手机用户) 联系我们: LSCA-China_800@agilent.com 在线询价: www.agilent.com/chem/erfq-cn www.agilent.com 仅限研究使用。不可用于诊断目的。 本文中的信息、说明和指标如有变更,恕不另行通知。 ◎安捷伦科技(中国)有限公司,2016 2016年5月6日, 中国出版 5991-6810CHCN Agilent Technologies 摘要在缺少动态信息的情况下,代谢组学数据的解读通常十分复杂,这是因为在相应代谢物丰度未发生改变时,某个通路的代谢流可能已经发生显著变化。稳定同位素示踪( 定性代谢流分析)的巨大潜力有助于解决这些状况,以便更深入地了解生物系统。在定性代谢流分析中,将稳定同位素示踪剂(通常含有 13C、15N 或 2H)引入生物系统,导致下游代谢物的同位素模式(同位素异数体分布)发生改变。该方法尤其适用于易于引入稳定同位素示踪剂的细胞模型,对产生的标记模式及示踪剂掺入动力学的分析将提供对酶功能、途径依赖、基因表达和蛋白功能变化效应的洞察。Agilent MassHunter VistaFlux 软件旨在设计一套完整的工作流程,帮助科学家进行定性代谢流分析,助于高分辨率准确质量数 TOF 和 Q-TOF LC/MS 数据的处理和可视化。软件提供的工具可进行特征提取、同位素掺入分析、同位素异数体丰度分析、天然丰度校正、统计学分析以及数据在通路中的可视化。本应用简报展示了使用 13C5 - 谷氨酰胺(U-13C-Gln) 作为示踪剂,对软骨肉瘤细胞系进行定性代谢流分析的研究示例。采用安捷伦 LC/TOF 系统进行数据采集,并使用 MassHunter VistaFlux 进行数据分析和解析。研究证明了异柠檬酸脱氢酶 2 (IDH2)的突变对三羧酸循环代谢产生的影响,特别是突变的 IDH2 异构体将致使谷氨酰胺转化生成 2-羟基戊二酸 (2-HG)。前言近期,多种实体瘤和血液肿瘤中均发现了体细胞 IDH 突变,其中包括胶质瘤、急性髓性白血病和软骨肉瘤。异柠檬酸脱氢酶(IDH1/2) 的突变发生在酶的活性位点上,形成的功能获得性表型导致代谢物 2-羟戊二酸 (2-HG) 的生成能力提高。近期研究表明,IDH 突变细胞中 2-HG 的高水平累积与清除途径的饱和相关,因此 2-HG 成为了 DNA 脱甲基酶 TET 家族、组氨酸脱甲基酶 Jumonji C 家族、mRNA 脱甲基酶 FTO 和 ALKBH5 以及调节缺氧诱导因子 (HIF) 的脯氨酰羟化酶 (PHD) 等各种 α-酮戊二酸 (α-KG)依赖型加双氧酶的竞争性抑制剂。这些变化促进了抑制性染色质环境的形成,这种环境可将细胞锁定在未分化状态下。软骨肉瘤是由软骨细胞沉积软骨引起的一种肉瘤,具有较高的 IDH 突变发生率。软骨肉瘤肿瘤来源细胞系的易得性使其成为了解 IDH 突变后果的实用模型系统。其中一项重要研究是使用定性代谢流分析来说明突变酶产生的 2-HG 来自 α-KG 和三羧酸循环中的谷氨酰胺。结论采用 Agilent MassHunter VistaFlux 工作流程,在 CS-1 软骨肉瘤细胞中以 U-13C-Gln 作为代谢示踪剂进行定性代谢流分析。该分析明确证实了 IDH2 突变异构体对谷氨酰胺衍生的 α-KG 到 2-HG 的转化活性。8 小时后,IDH 突变细胞中大部分 2-HG 组均被标记为 M+5 同位素异数体。IDH2 R172S 突变酶的新生变形活性超过了内源性 2-HG 脱氢酶,因此 2-HG 的生成量有所增加,从而使 2-HG 的累积浓度升高。因此,2-HG 成为了 DNA 脱甲基酶 TET 家族和组氨酸脱甲基酶 Jumonji C 家族等多种 α-KG 依赖型加双氧酶的竞争抑制剂,造成了包括 DNA 超甲基化在内的表观遗传学变化,促进了未分化染色质环境的产生。分析还展示了 U-13C-Gln 示踪剂进入经典三羧酸循环通路中的结果。与手动数据挖掘相比,VistaFlux 工作流程为稳定同位素示踪数据处理提供了全面、自动化且快捷的框架,其中包括同位素异数体提取和定性代谢流文件数据的通路可视化。
确定






还剩8页未读,是否继续阅读?

产品配置单


安捷伦科技(中国)有限公司为您提供《软骨肉瘤细胞系中13C-谷氨酰胺检测方案(自动进样器)》,该方案主要用于癌细胞/肿瘤细胞中13C-谷氨酰胺检测,参考标准--,《软骨肉瘤细胞系中13C-谷氨酰胺检测方案(自动进样器)》用到的仪器有Agilent 1290 Infinity II Multisampler、Agilent 1290 Infinity II 液相色谱系统
推荐专场






相关方案
更多
该厂商其他方案
更多