








方案详情
文
以寒鸦为测试对象。在风洞中,采用LaVision公司的时间分辨2D3C立体PIV测量系统,对开槽翼尖的多核漩涡对鸟类滑翔和扑翼飞行的支撑作用进行了研究。实验装置包括一台英国Litron公司的LDY304PIV高重复频率激光器,两台Imager pro HS 4M型高速相机,和DaVis8.4软件平台构成。
方案详情

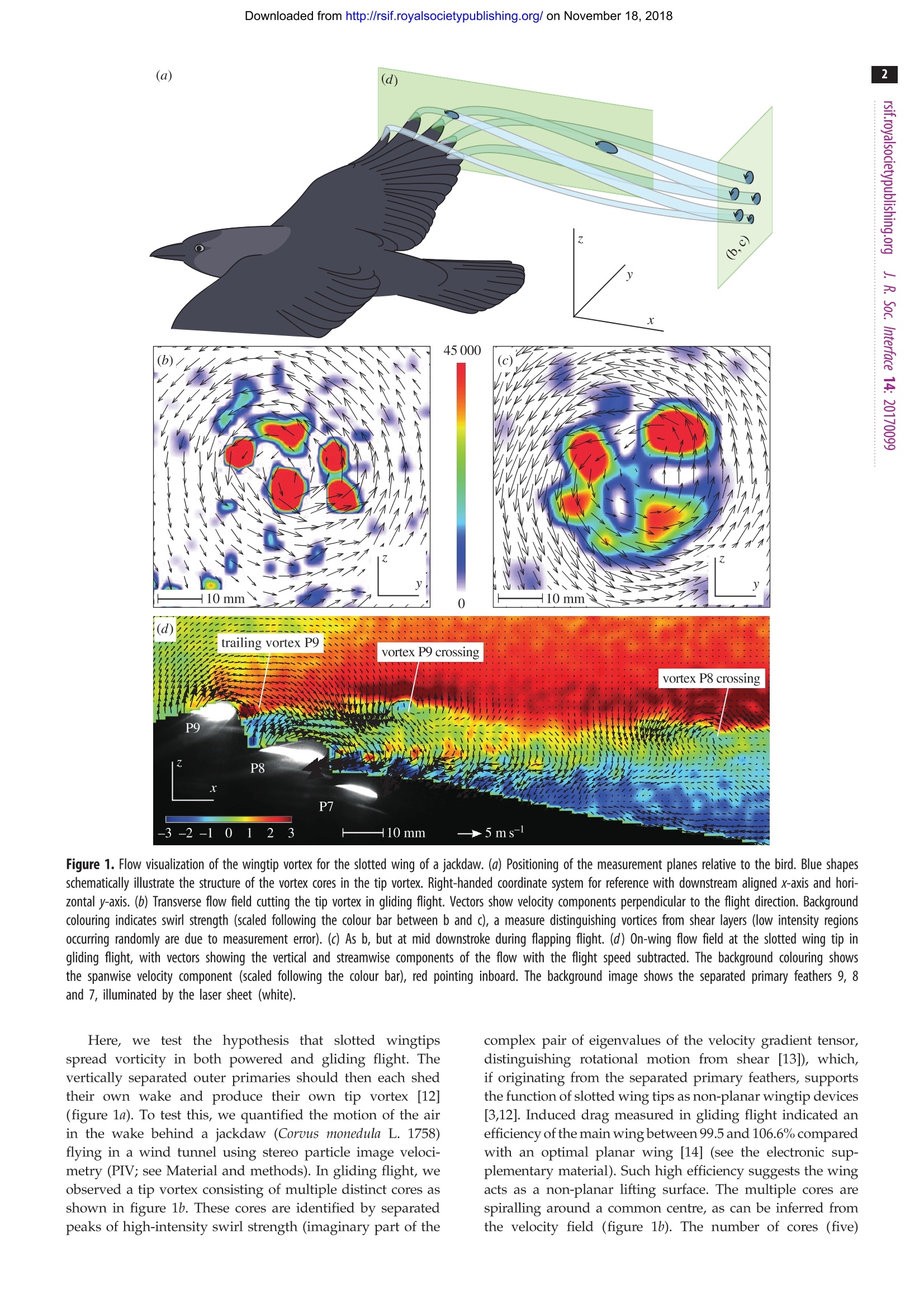
Downloaded from http://rsif.royalsocietypublishing.org/ on November 18, 2018 THE ROYAL SOCIETYPUBLISHINGC 2017 The Author(s) Published by the Royal Society. All rights reserved. INTERFACE rsif.royalsocietypublishing.org ReportCheck for updates Cite this article: KleinHeerenbrink M, Johansson LC, Hedenstrom A. 2017 Multi-coredvortices support function of slotted wing tipsof birds in gliding and flapping flight. . R. Soc.Interface 14: 20170099. http://dx.doi.org/10.1098/rsif.2017.0099 Keywords: Author for correspondence:Marco KleinHeerenbrinke-mail: marco.kleinheerenbrink@zoo.ox.ac.uk Electronic supplementary material is availableonline at https://dx.doi.org/10.6084/m9.figshare.c.3773297. Multi-cored vortices support functionof slotted wing tips of birds in glidingand flapping flight Marco KleinHeerenbrink12, L. Christoffer Johansson1 and Anders Hedenstrom1 Department of Biology, Lund University, Lund, Sweden Department of Zoology, University of Oxford, Oxford, UK iDMK, 0000-0001-8172-8121;LO, 0000-0002-1851-3635; AH, 0000-0002-1757-0945 Slotted wing tips of birds are commonly considered an adaptation to improvesoaring performance, despite their presence in species that neither soar norglide. We used particle image velocimetry to measure the airflow aroundthe slotted wing tip of a jackdaw (Corous monedula) as well as in its wakeduring unrestrained flight in a wind tunnel. The separated primary feathersproduce individual wakes, confirming a multi-slotted function, in both glidingand flapping flight. The resulting multi-cored wingtip vortex represents aspreading of vorticity, which has previously been suggested as indicative ofincreased aerodynamic efficiency. Considering benefits of the slotted wingtips that are specific to flapping flight combined with the wide phylogeneticoccurrence of this configuration, we propose the hypothesis that slottedwings evolved initially to improve performance in powered flight. 1. Introduction Slotted wing tips, where the outer primary feathers split and spread vertically,are found in a wide range of birds and are considered an adaptation for efficientflight [1,2]. Pronounced slotted configurations are facilitated by emargina-tions on the vanes of the outer primary feathers. The broader bases of thefeathers overlap, limiting twisting and bending of individual feathers, whilethe narrower outer parts may deform independently of each other. Underaerodynamic loads, the tips of the leading primaries rise above those of thetrailing feathers, forming a staggered, non-planar, configuration. Previousresearch has focused particularly on the benefits of the vertical separation ofthe outer primaries in thermal soaring birds [2-4], where wingtip slots werefound to reduce induced drag (the cost of producing lift with finite wings,also referred to as vortex drag [5]). For a planar wing, induced drag is mini-mized by an elliptical lift distribution that comes with a spanwise uniformdownwash distribution. Non-planar configurations can spread vorticity verti-cally, abating high induced flows near the tip of a more uniformly loadedwing, allowing for reduced drag for a given wingspan [2,6,7] (see the electronicsupplementary material). Extensive research into this subject since the earlytwentieth century, has led to the development of wingtip devices that arenow common in passenger aircraft [7]. Many bird species that specialize in gliding flight do not exhibit a slottedwingtip configuration, and have instead evolved high aspect ratio wings, analternative solution to reduce induced drag, by extending their leading primariesto form a pointed wing, as seen in, for example, petrels and albatrosses (Procellar-iiformes) [8]. On the other hand, feather emarginations, indicating a slottedconfiguration, are found in many bird species that neither soar nor glide [9,10],across the avian phylogenetic tree [8,11] and among the most basal lineagesof birds, including species that only fly occasionally (e.g. Tinamiformes,Galliformes). This suggests a more general function and benefit of a slottedwing tip than for gliding flight only. Figure 1. Flow visualization of the wingtip vortex for the slotted wing of a jackdaw. (a) Positioning of the measurement planes relative to the bird. Blue shapesschematically illustrate the structure of the vortex cores in the tip vortex. Right-handed coordinate system for reference with downstream aligned x-axis and hori-zontal y-axis. (b) Transverse flow field cutting the tip vortex in gliding flight. Vectors show velocity components perpendicular to the flight direction. Backgroundcolouring indicates swirl strength (scaled following the colour bar between b and c), a measure distinguishing vortices from shear layers (low intensity regionsoccurring randomly are due to measurement error). (c) As b, but at mid downstroke during flapping flight. (d) On-wing flow field at the slotted wing tip ingliding flight, with vectors showing the vertical and streamwise components of the flow with the flight speed subtracted. The background colouring showsthe spanwise velocity component (scaled following the colour bar), red pointing inboard. The background image shows the separated primary feathers 9, 8and 7, illuminated by the laser sheet (white). Here, we test the hypothesis that slotted wingtipsspread vorticity in both powered and gliding flight. Thevertically separated outer primaries should then each shedtheir own wake and produce their own tip vortex [12](figure 1a). To test this, we quantified the motion of the airin the wake behind a jackdaw (Corous monedula L. 1758)flying in a wind tunnel using stereo particle imageveloci-metry (PIV; see Material and methods).In gliding flight, weobserved a tip vortex consisting of multiple distinct cores asshown in figure 1b. These cores are identified by separatedpeaks of high-intensity swirl strength (imaginary part of the complex pair of eigenvalues of the velocity gradient tensor,distinguishing rotational motion from shear [13]), which,if originating from the separated primary feathers, supportsthe function of slotted wing tips as non-planar wingtip devices[3,12]. Induced drag measured in gliding flight indicated anefficiency of the main wing between 99.5 and 106.6% comparedwith an optimal planar wing [14] (see the electronic sup-plementary material). Such high efficiency suggests the wingacts as a non-planar lifting surface. The multiple cores arespiralling around a common centre, as can be inferred fromthe velocity field (figure 1b). The number of cores (five) in the wingtip vortex matches the number of slotted feathersof the wing tip. The wake shed during flapping flight showed a similarwingtip vortex structure to that found in gliding flight, com-posed of multiple cores (figure 1c), demonstrating that alsoduring powered flight the separated feathers spread the vor-ticity of the tip vortex. This implies that an induced dragreduction may be realized in both flapping flight and glidingflight. In flapping flight, the force and circulation distributionvary throughout the wingbeat, which negatively affects thespan efficiency [15,16]. This makes comparison to an ideal-ized optimally loaded un-slotted wing in flapping flightchallenging and beyond the scope of this paper. To determine whether the multiple cores we found in the tipvortex originated from the individual feathers, we measured thevelocity field on top of the wing in a streamwise plane, close tothe wing tip, where the feathers separate. The flow field is visual-ized in figure 1d, with the spanwise flow (out of plane) indicatedby background colouring and the in-plane velocity (with flightspeed subtracted) by vectors. A streamwise vortex comingfrom the tip of the ninth primary (P9) can be identified by thespanwise velocity component: an inward flow (positive) can beseen on top,and outward flow (negative) below the core, demon-strating the rotation of the P9 tip vortex. About 40 mmdownstream of P9, this vortex crosses the measurement plane,indicated by the strong (in-plane) counterclockwise rotation, asit is convected inboard by the overall counterclockwise rotationof the wingtip vortex. The tip of the following feather (P8) isoutboard of our measurement plane. Approximately, 90 mmdownstream of P8 its tip vortex crosses the measurement plane,identified by the strong (in-plane) counterclockwise rotation.The cores rotate at approximately 15 mm from the overall wing-tip vortex centre (figure 1b), with a tangential speed of about3 ms(figure 1d). At a flight speed of 8.5 ms, the coreshave a wavelength of approximately 270 mm. This means thatthe vortex from P8 has travelled 1/3 cycle when it crosses themeasurement plane, which agrees with the vertical location ofthe crossing. From these observations, we conclude that the mul-tiple cores of the tip vortices, found in the wakes of the glidingand flapping bird, originate from the separated primary feathers. Besides reducing induced drag, slotted wing tips may haveadditional aerodynamic benefits specific to flapping flight.During the downstroke, the air approaches the wing tip at ahigh angle of attack, which would make the distal part of thewing prone to stall. To reduce the angle of attack, the wingtip needs to twist (washout), like the blade of a propeller.Instead of twisting the entire wing, which would require struc-turally challenging bending of the feather shafts, slottingallows feathers to twist individually [1]. This allows for abroader wing tip, and thus a higher loading of the outboardwing, which is most efficient for thrust production [17] andas such beneficial for powered flight. Additionally, a multi-slotted wing can increase the maximum lift coefficient of anaerofoil [18], thereby maximizing the available thrust. Beingable to generate high thrust is beneficial,e.g. for predator eva-sive manoeuvr1esr, including take-off in ground living species,and would, therefore, be highly selected for. In this study, we demonstrate that slotted wing tips causevortex spreading-the mechanism behind reduced induceddrag-in both gliding and flapping flight of a jackdaw. In thelight of additional aerodynamic advantages unique to flappingflight (e.g. reduced risk of stall and increased thrust), andthe abundance of slotted wing configurations across the phylogeny of modern birds, we propose the hypothesis thatthe slotted wing tip originally evolved to improve flappingflight performance. Testing this hypothesis requires a widecomparative analysis of feather morphology and flight styleand further studies of the aerodynamic details of slottedwing tips during flapping flight. 2. Material and methods 2.1. Study species A young jackdaw was taken from a study colony near Revingehed,Skane, Sweden (11 June 2013), and trained to keep position whenflying in the wind tunnel. For more details, see [14]. FollowingPennycuick et al.[19], maximum wingspan was measured fromthe bird in the hand, being bw=0.67 m, and wing area was deter-mined from a tracing of the wing: Sw=0.0652 m² (including bothwings and the area between the wing roots). Body mass wasmeasured daily and was stable around 0.215 kg. 2.2. Experimental set-up We used the low-turbulence tiltable wind tunnel at the Departmentof Biology, Lund University, Sweden [20], which has an octagonaltest section with a width of 1.2 mand a height of 1.08 m. For glidingflight experiments, the tunnel was set at a glide angle of 6°. Forpowered flight, the wind tunnel was horizontal. In all reported-1cases, the airspeed was in the range of 8-8.5 ms. The velocity field in a plane perpendicular to the free streamflow in the wake behind the bird was measured using a stereoPIV system recording 640 frame-pairs per second. A 527 nmdiode pumped LDY304PIV laser (Litron Lasers Ltd, Rugby, UK)was used to illuminate particles (approx. 1 um) in a sheet(approx. 3 mm thick). For wake experiments, two LaVisionImager pro HS 4M (LaVision Gmbh, Gottingen, Germany)high-speed cameras 2016x2016 px in stereo configuration wereaimed at the left wingtip vortex (note: shown mirrored horizon-tally in figure 1). Analyses were performed in Davis 8.3(LaVision Gmbh), with a sum of correlation routine on a singleimage pair, using final box size 24×24 px with 50% overlap.The resolved flow field had a resolution of 6 vectors cmforgliding and 3 vectors cm-for the flapping flight set-up. On-wing experiments were performed while the bird wasgliding. We used the same PIV system as above, but with thefield of view parallel to the sagittal plane. The laser sheet wasprojected from downstream of the bird at a shallow angle fromabove with respect to the wind tunnel axis. The cameras werepositioned on the opposite side of the bird to the wing studied,making a shallow angle with the wing surface. The images wereanalysed using 16 x16 px boxes with 50% overlap generating avector resolution of 13.3 vectors cm Ethics. The study was performed in accordance with approved exper-imental guidelines. Procedures were approved by the Malmo-Lundanimal ethics committee (M 33-13). ( D ata accessibili t y. The vector dat a shown i n figure 1 h ave been up l oaded in t ab-separated format as t h e electronic supplementary mat e rial. ) ( Authors contribut i on s. M.K. and L.C. J . conceive d th e study ; M.K. trainedthe bird;M.K. and L.C.J. carried out the experiments; M.K., L . C.J. andA.H. d rafted the manuscript. ) ( Com p eti n g interest s. We declare we have no competing interests. ) ( Funding . The researc h receive d funding from the Swedish ResearchCouncil t o A .H. ( 6 21-2009-4965/621-2012-3585) and to L. C .J. (621-2013-4596), from an i n frastructure grant f rom L und University toA.H. a nd from t h e R o yal P h ysiographic Society i n L u nd to M. K .This work r eceived support f rom t he Centre f or Animal M ovementResearch ( CAnMove) fi n anced by a Linn a eus gra n t (34 9 -2007-8690) from the Swedish Research Council and Lund Un i versity. ) Acknowledgements. We thank Johan Nilsson for providing the jackdaw.We are also grateful to the other members of the Animal Flight Laboratory for their occasional assistance during the experimentsand bird care. ( 1. Withers PC. 1981 The aerodynamic performance ofthe w ing in R e d-shouldered H aw k Buteo linearisand a p o ssible a e roelastic role o f wing-tip slots. l b is(Lond. 1859) 123,239-247.(doi:10.1111/j.1474- 919X.1981.tb00933.x) ) 2. Tucker VA. 1993 Gliding birds: reduction of induceddrag by wing tip slots between the primaryfeathers. J. Exp. Biol. 180, 285-310. 3. Tucker VA. 1995 Drag reduction by wing tip slots ina gliding Harris’ hawk, Parabuteo unicinctus. J. Exp.Biol. 198, 775-781. 4. Eder H, Fiedler W, Neuhauser M. 2015 Evaluation ofaerodynamic parameters from infrared laser trackingof free-gliding white storks. J. Ornithol. 156,667-677. (doi:10.1007/s10336-015-1176-7) 5. Anderson JDJ. 2007 Fundamentals of aerodynamics,4th edn. New York, NY: McGraw-Hill Education. 6. Prandtl L. 1924 Induced drag of multiplanes.US Patent NACA-Tn-18. 7. Kroo IM. 2001 Drag due to lift: concepts forprediction. Annu. Rev. Fluid Mech. 33, 587-617.(doi:10.1146/annurev.fluid.33.1.587) ( 8. Feo TJ, Field DJ , Prum RO. 2015 Barb geometryof a symmetrical fe a thers reveals a t r a n sitional m orphology in the evo l ution of a vian flight. ) Proc. R. Soc. B 282, 20142864. (doi:10.1098/rspb.2014.2864) 9. Savile DBO. 1957 Adaptive evolution in the avianwing. Evolution (NY) 11, 212-224. ( 10. Norberg RA. 1985 Function of vane asymmetry and s haft curvature in bird fl i ght feathers: inf e rences o n flight ability of a rchaeopteryx. In Th e beg i nningsof birds (ed.MK Hecht), pp. 303-318.Eichstatt, Germany: Freunde d es J u ra-Museums Eichstatt. ) ( 11. Prum R O , B e rv J S , Dornburg A , Field D J, TownsendJP, Lemmon E M, L emmon A R . 2 0 15 A c omprehensive phylogeny o f birds ( Aves)using targeted next-generation DNA sequencing.Nature 5 26, 569-573.(d o i:10.1038/nature15697) ) ( 12 . Co iro D P , Nicolosi F , S c herillo F, Maisto U. 20 0 8 S ingle v ersus multiple winglets: numerical and e xperimental investigation. I n 2 6 th Int. CongressAeronaut. Sci . Anchorage , AK , 14- 1 9 September, pp. 1- 1 1. ) ( 13 . Z hou J , Adrian RJ, Balachandran S , K e ndall TM. 1999 Mechanisms for generating coherent p ackets o f h a irpin v o rtices in c ha nnel flow. J. Fluid Mech. 387, 353-396. ( doi:10.1017/ S002211209900467X) ) 14. KKleinHeerenbrink M, Warfvinge K, Hedenstrom A.2016 Wake analysis of aerodynamic components forthe glide envelope of a jackdaw (Corvus monedula).J. Exp. Biol. 219, 1572-1581. (doi:10.1242/jeb.132480) 15. SSachs G. 2015 Aerodynamic cost of flapping.J. Bionic Eng. 12, 61-69.(doi:10.1016/S1672-6529(14)60100-1) ( 16. K K lein Heerenbrink M, Johansson L C, Hedenstrom A. 2015 P ower of t he wingbeat: modelling the e ffects of f lapping wings i n vertebrate flight. Proc. R. Soc. A 471, 2 0140952. (doi:10.1098/rspa. 2014.0952) ) ( 17. Hall K C, Hall SR. 1996 Minimum induced p ower r e quirements f o r f l apping flight. J. FluidMech. 323 , 285-315. ( doi:10.1017/S002211 2096000924) ) ( 18 . . S mith AMO. 1975 Hig h -lift aerodynamics. J. Aircr . 12 , 501-530. (doi : 10.2514/3.59830) ) ( 19.. P P ennycuick C J . 2008 Modelling the fl y ing bi r d. A msterdam, The Netherlands: Elsevier. ) ( 20. P P ennycuick C J , A lerstam T , H edenstrom A. 19 9 7A n ew l o w-turbulence wind t u nnel f o r bird f light e xperiments at L u nd Uni v ersity, Sweden. J. E x p . Biol . 200,1441-1449. ) Slotted wing tips of birds are commonly considered an adaptation to improve soaring performance, despite their presence in species that neither soar nor glide. We used particle image velocimetry to measure the airflow around the slotted wing tip of a jackdaw (Corvus monedula) as well as in its wake during unrestrained flight in a wind tunnel. The separated primary feathers produce individual wakes, confirming a multi-slotted function, in both glidingand flapping flight. The resulting multi-cored wingtip vortex represents a spreading of vorticity, which has previously been suggested as indicative of increased aerodynamic efficiency. Considering benefits of the slotted wing tips that are specific to flapping flight combined with the wide phylogenetic occurrence of this configuration, we propose the hypothesis that slottedwings evolved initially to improve performance in powered flight.
确定



还剩2页未读,是否继续阅读?

产品配置单


北京欧兰科技发展有限公司为您提供《开槽翼尖中速度矢量场检测方案(粒子图像测速)》,该方案主要用于航空中速度矢量场检测,参考标准--,《开槽翼尖中速度矢量场检测方案(粒子图像测速)》用到的仪器有三维立体PIV、LaVision DaVis 智能成像软件平台
推荐专场






相关方案
更多
该厂商其他方案
更多