








厌氧真菌培养上清液或混合瘤胃饲料作为新型青贮添加剂的适宜性Suitability of anaerobic fungi culture supernatant or mixed ruminal fluid as novel silage additives
方案详情

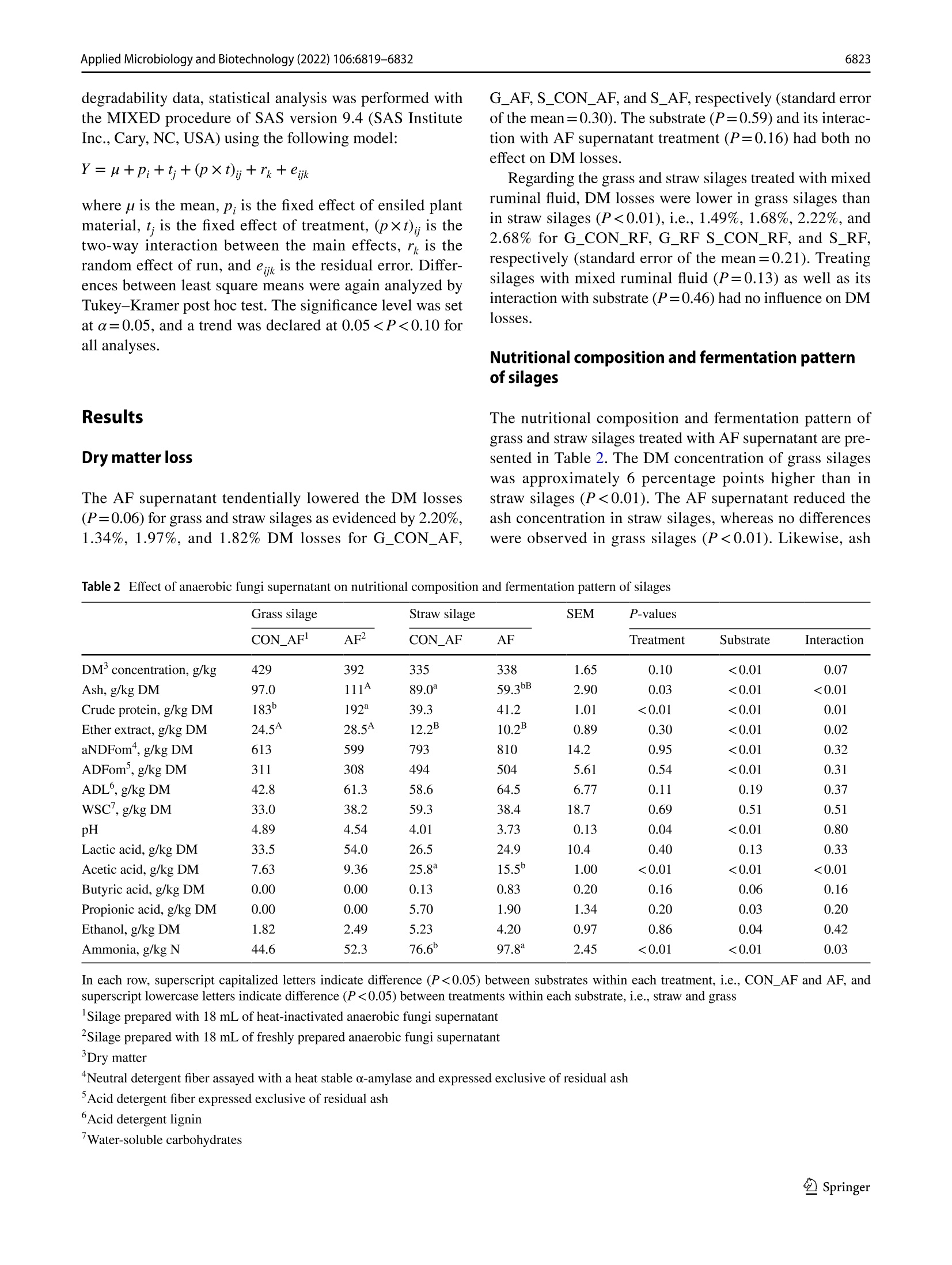
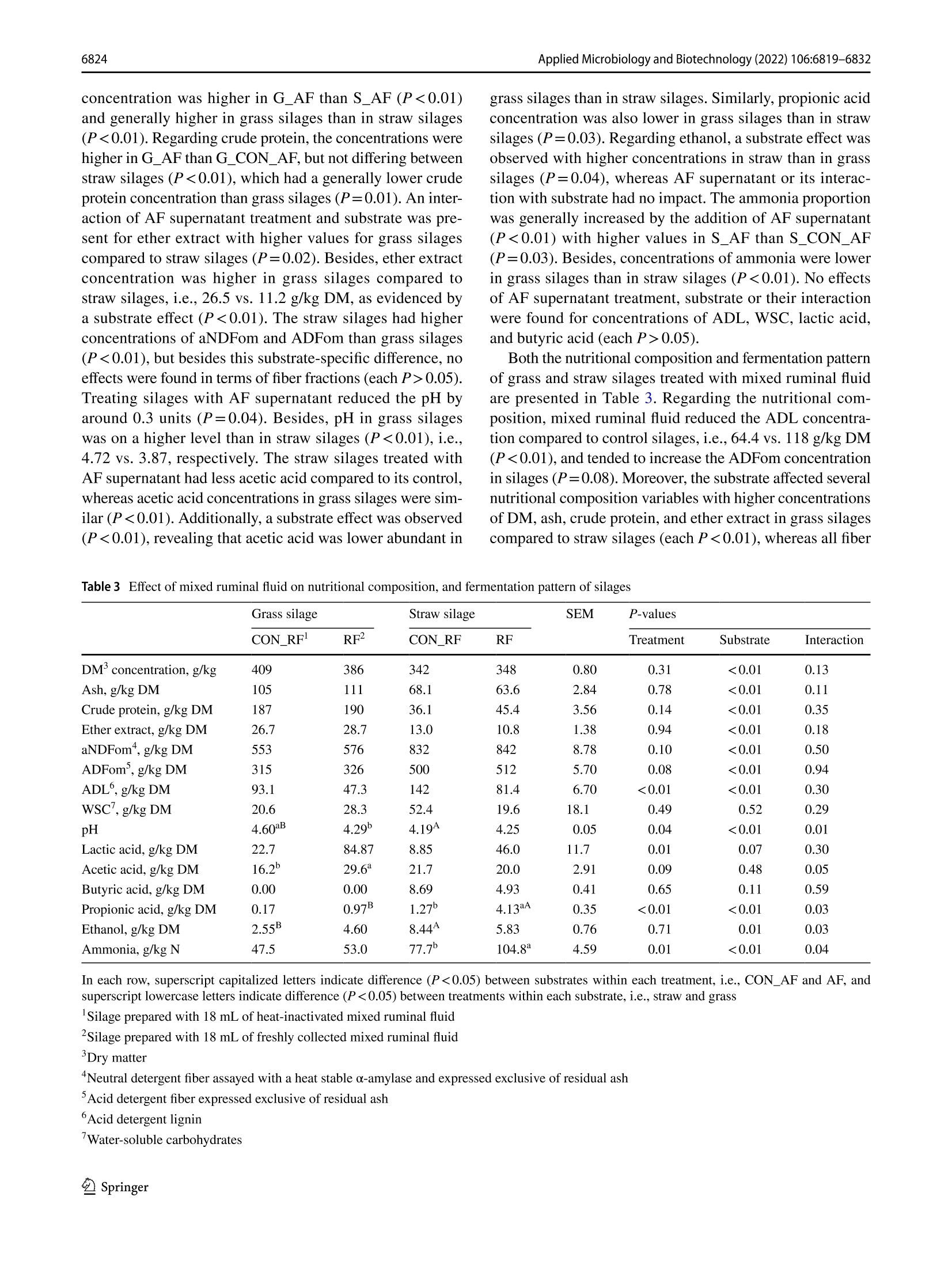
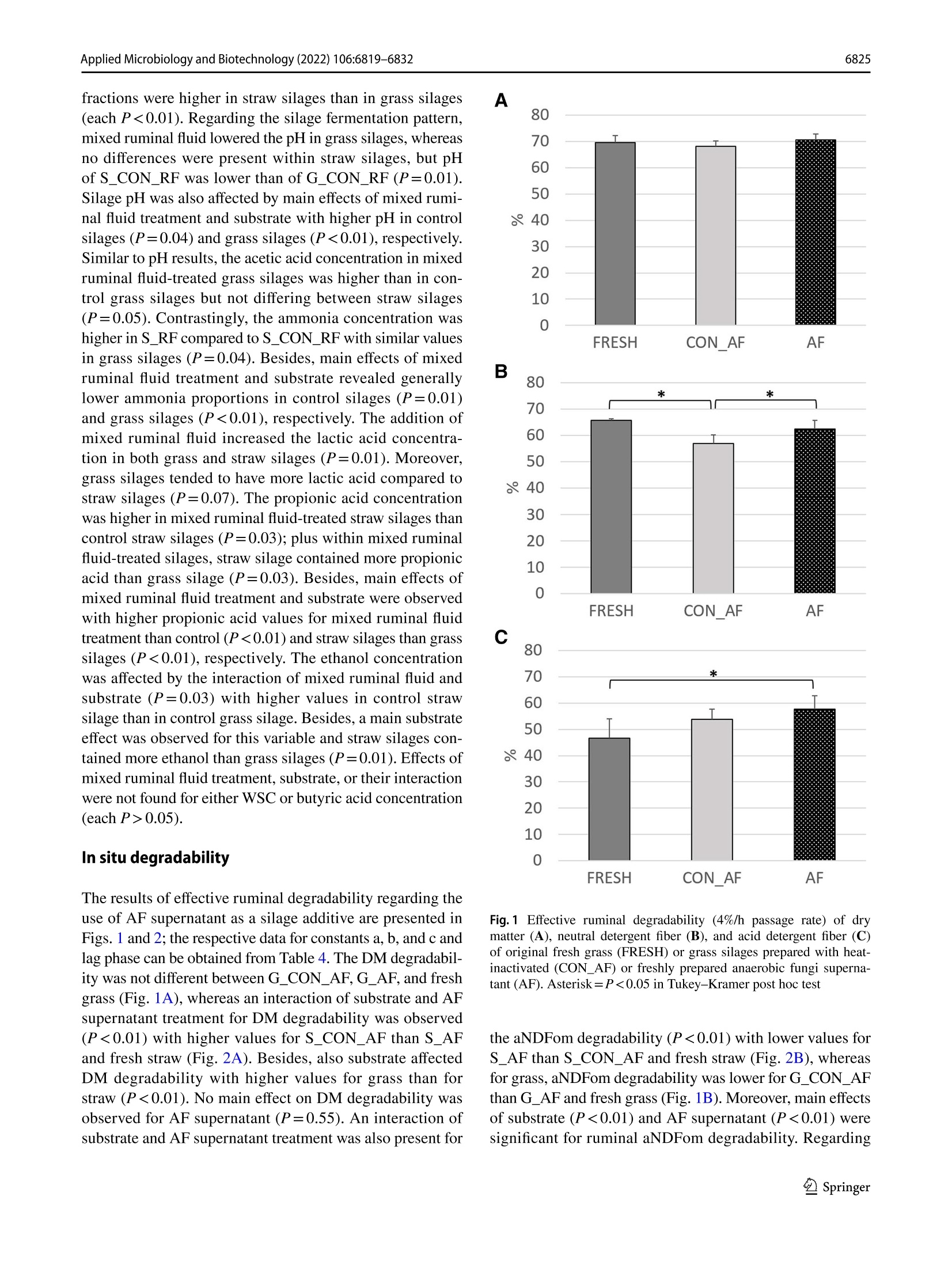
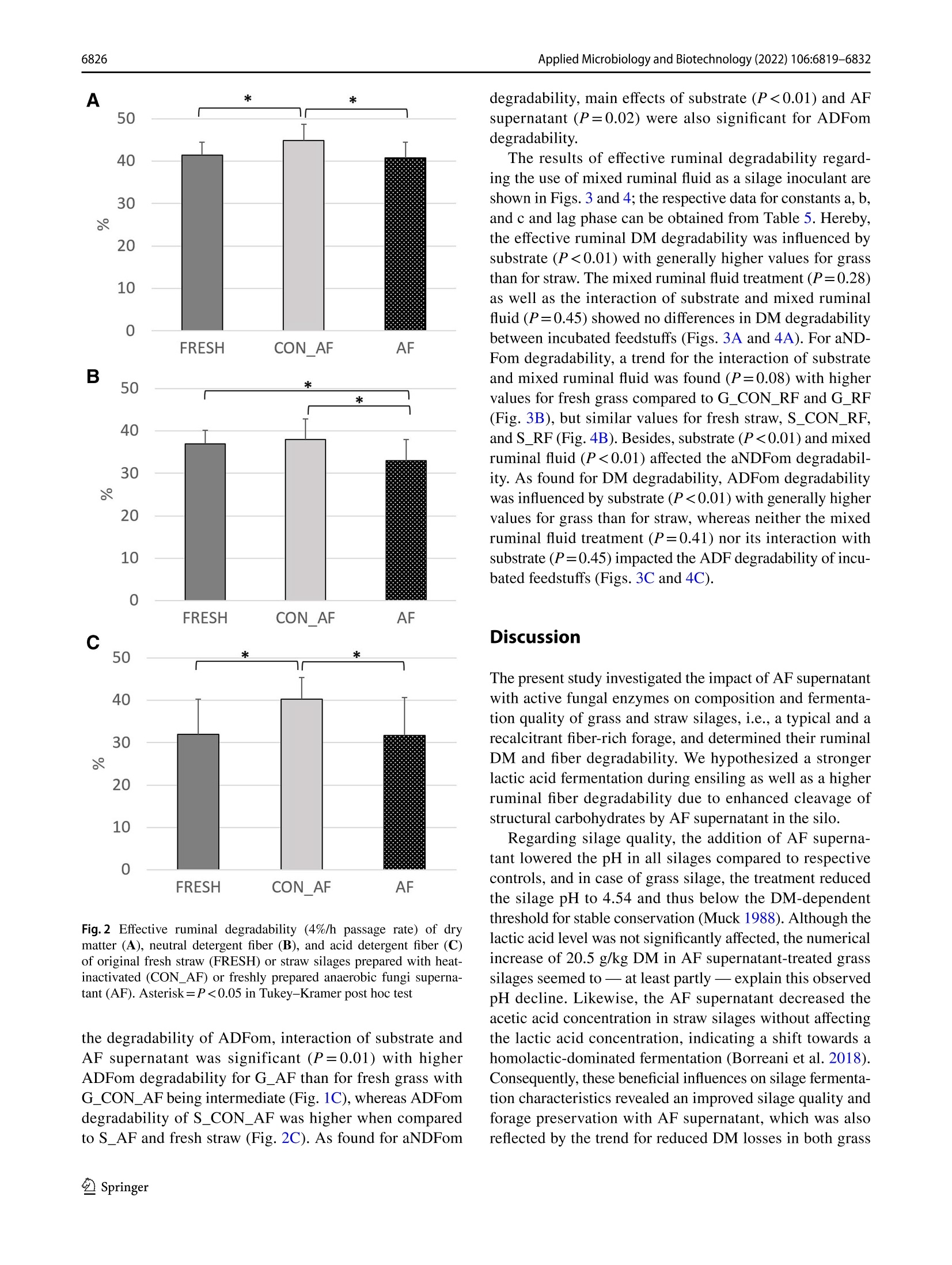
厌氧真菌培养上清液或混合瘤胃饲料作为新型青贮添加剂的适宜性Suitability of anaerobic fungi culture supernatant or mixed ruminal fluid as novel silage additivesApplied Microbiology and Biotechnology (2022) 106:6819-6832https://doi.org/10.1007/s00253-022-12157-WENVIRONMENTAL BIOTECHNOLOGY Applied Microbiology and Biotechnology (2022) 106:6819-68326820 厌氧真菌培养上清液或混合瘤胃饲料作为新型青贮添加剂的适宜性 Suitability of anaerobic fungi culture supernatant or mixed ruminal fluid as novel silage additives Thomas Hartinger D . Katerina Fliegerova²D.Qendrim Zebeli11D Received: 6 June 2022 / Revised: 12 August 2022/ Accepted: 24 August 2022 /Published online: 14 September 2022O The Author(s) 2022 Abstract This study investigated silage quality characteristics and ruminal f iber degradability of grass and straw ensiled with either anaerobic fungi (AF) supernatant with active fungal enzymes or mixed ruminal fluid as novel silage additives. Compared to control silages, AF supernatant improved the quality of grass and straw silages as evidenced by decreased pH, acetic acid concentration, and dry matter losses. Likewise, mixed ruminal fluid enhanced lactic acid fermentation, which f urther resulted in lower pH of the treated grass silage. The ruminal fiber degradability was determined using in situ incubations and, com-pared to controls, the cellulose degradability was higher for grass silage with AF supernatant, whereas ruminal degradability of straw silage was reduced by this treatment . In contrast , mixed ruminal f l uid did not influence f iber degradability of silages in the rumen. Concluding, both novel additives improved silage quality, whereas only AF supernatant enhanced ruminal fiber degradabil i ty of grass silage and therefore may represent an approach for improving forage utilization by ruminants. Key points ·Enzymes ofanaerobic fungi supernatant improve quality of grass and straw silages. · Mixed ruminal f luid enhances lactic acid fermentation when ensiling grass and straw. ·Enzymes ofanaerobic fungi supernatant increase ruminal grass silage degradability. Keywords Anaerobic fungi· Enzymes · Forage · Ruminant· S ilage additive From physiological, ecological , and economical perspec-tives, silages constitute the main dietary component in cattle feeding, providing energy, nutrients, and struc t ural fiber. An excellent silage quality with high ruminal degradabi l ity is of utmost importance to adequately meet the cat t le’s nutritive demands for performance while decreasing the inclusion of concentrates in the diet. However, various abiotic and biotic factors may often cause poor forage quality that is character-ized by fewer sugars but more recalcitrant fibrous structures, Thomas Har t inger thomas.hartinger@vetmeduni .ac.at 1 Institute of Animal Nutrition and Functional Plant Compounds, Department for Farm Animals and Veterinary Publ i c Health , University of Veterinary Medicine Vienna,Veterinarplatz 1, 1210 Vienna, Austria 2 Laboratory of Anaerobic Microbiology, Institute of Animal Physiology and Genetics, Czech Academy of Sciences,14220 Prague, Czech Republic which eventually reduce the nutritive value and ruminal deg-radability of these forages (Lauer 2012; Borreani et al . 2018)and therefore jeopardize the adequate energy and nutrient exploitation by the animal . In thi s context , the future trend towards higher aridity in Europe (Forzieri et al . 2014) will increase the risk of drought-impaired and thus low-quality forage, which in turn emphasize the necessity for develop-ing strategies that ensure the sufficient energy supply from s i lages of different substrate quali t ies. Remarkably, the ensi l ing process i tself has been dem-onstrated to slightly improve fiber degradability by decon-struc t ing l ignocellulosic complexes (Ambye-Jensen et al.2013; Zhao et al. 2018). The use of novel silage additives based on enzymes originating from anaerobic fungi (AF)culture medium has been recently suggested as a way to more effectively pre-cleave fibrous structures in silages (Har t inger and Zebeli 2021). Anaerobic fungi are com-mensals of the rumen microbiome and ef f iciently degrade fiber using various enzymes as well as physical penetra-tion via fungal hyphae (Hess et al . 2020). So far, only few studies investigated the use of viabl e fungal cultures during ensi l ing of r i ce straw and whole-crop corn. Thereby, treat-ment with viable fungal cultures resulted in enhanced lactic acid fermentation as well as higher ruminal degradability for both substrates (Lee et al. 2015; Wang et al . 2019). How-ever, there is credible evidence that those improvements in AF-treated silages were not related to the viable AF cul-tures as they cannot survive in the silo (Lee et al . 2015),therefore rather suggesting no physical penetration of plant part i cles by fungal hyphae. More l ikely, the extracellularly fungal enzymes present in the culture medium (Haitjema et al. 2014.2017) caused those beneficial effects observed (Hartinger and Zebel i 2021). Due to their wide pH range and affinity for recalcitrant f iber components (Hagen et al.2020; Wang et al. 2019), AF enzymes may indeed pre-cleave f ibrous compounds during ensiling and subsequently allow an enhanced degradability of silages in the rumen.Besides, activity of AF enzymes can release glucose from fiber breakdown (Wang et al. 2011), which in consequence could support lactic acid fermentation i n the silo and thereby improve substrate conservation. Therefore, the first aim of our study was to investigate the effects of the addition of AF supernatant with fungal enzymes on the composition and in situ fiber degradability of grass or wheat straw silages,i.e., a common forage in cattle feeding and a by-product of grain production that is very rich in recalcitrant f iber.We hypothesized a stronger lactic acid fermentation in the silos as well as a higher in situ degradability due to activity of fungal enzymes when AF supernatant is included as a silage additive. Since the isolation and cultivation of AF are complex and laborious (Dollhofer et al . 2015), a direct inoculation with ruminal f luid, which includes the rumen microbiome's diverse enzymatic repertoire and consequently as well AF enzymes (Puniya et al. 2015), might constitute an analo-gous approach to tackle plant fiber structures already during ensiling. Up to now, ruminal fluid has not been explored as a silage additive , but in fact represents a reservoir of fibrolytics and their enzymatic tools, which may be ben-eficially used for ensiling. Therefore, a second aim of the study was to evaluate the suitabi l ity of mixed ruminal fluid as an inoculant for grass and straw silages. Compared to control silages, both a higher lactic acid fermentation and in si t u degradabi l ity of silages t reated with mixed ruminal fluid were expected. Materials and methods Preparation of anaerobic fungi supernatant as silage additive For receiving the AF enzymes-based additive, i .e., t h e AF supernatant, an AF mixture isolated from the ovine rumen was anaerobically cultivated at 39 °℃ on M10 medium (Caldwell and Bryant 1966) enriched by 25% (v/v) rumen fluid with 4 g/l of xylan (Serva Electrophoresis GmbH,Heidelberg, Germany) as carbon source. The AF mixture was then incubated in 100-ml bottles for 4 days, mycelia were collected by centrifugation at 4000×g for 20 min at 4C, and culture broth, i.e., AF supernatant, was further processed. The AF supernatant was tenfold concentrated by ultraf i l t ration (EMD Millipore 5121 Amicon St i rred Cell Model 8010, Cole-Parmer, Vernon Hills, IL , USA) under helium pressure of 300 kPa using membrane disc f i lters (B10K 76MM HIGH FLUX 10PK,Merck, Darmstadt, Ger-many) and subsequently lyophilized (Lyovac GT2, Leybold Heraeus, Cologne, Germany). The lyophi l ized powder was stored at -20 °℃ until i ts appl i cation. The eligibi l ity of AF supernatant was evaluated by determination of endo-1,4-β-xylanase (EC 3.2.1.8) and β-D-glucoside glucohydro-lase (E.C.3.2.1.21) activity according to methods of Lever (1977) and Bidochka et al. (1993), respectively. The enzy-matic activities of β-D-glucoside glucohydrolase and endo-1,4-B-xylanase were confirmed to be 261.8 nkat (170 ug of glucose/ml /h) and 1567.3 ukat (4,120 pg of xylose/ml/h),respectively. For the strain-speci f ic identi f ication of present AF, DNA from the cultivated mycelium was isolated using DNeasy PowerSoil Kit (Qiagen, Hilden, Germany) and the whole ITS region was ampl i fied using primer pair ITS1F (CTT GGTCATTTAGAGGAAGTAA; Gardes and Bruns 1993)and ITS4R (TCCTCCGCTTATTGATATGC; White et al.1990), resul t ing in amplicons of approximately 740 bp length. The AF identification was based on clone library approach as described previously by Mura et al. (2019).Briefly, plasmid DNA was isolated from 30 randomly selected ITS clones of Escherichia coli using a GenEluteTM HP Plasmid Miniprep Kit (Sigma-Aldrich, Burlington, MA,USA) and sent for Sanger sequencing using the M13F pr i m-ing sites within the vector pCR4 (SEQme, Dobris, Czech Republic). The sequences obtained from the cultivated AF mixture had highest similarity with three species of fun-gal genus Neocallimastix, i .e., Neocallimastix sp. NYR4(GenBank accession numbe r JQ782548, sequence identity 99.18%), Neocallimastix sp. WI3-B (GenBank accession number MK397979, sequence ident i ty 97.57%), and Neocal-limastix sp. NYF3 (GenBank accession number JQ782544,sequence identity 97.49%). Preparation of mixed ruminal fluid as silage additive The mixed ruminal fluid was obtained from a rumen-cannu-lated dry Holstein cow fed grass silage and grass hay (70:30,dry matter (DM) basis) ad l ibitum and additionally 1 kg of concentrate mixture (i .e., ground corn, barley, and wheat in a ratio of 0.5:0.25:0.25) per day that was provided in two equal portions in the morning and afternoon. The cow was fed and kept according to the Austrian guidelines of animal welfare (BGBl. I I Nr.485/2004 idF BGBl. I I Nr. 151/2017). Directly before ensiling, solid ruminal digesta was collected f rom the middle of the fiber mat and squeezed through three layers of gauze (Wilhelm Weisweiler GmbH & Co. KG, Miinster,Germany) to obtain particle-associated mixed ruminal fluid that was directly applied to the respective silages. Preparation of straw and grass silages For the experiment, silages were produced in 0.85-L glass jars (Weck GmbH u. Co. KG, Wehr, Germany) using either wilted grass [G] or wheat straw [S] as ensiling substrates.The chemical compositions of both substrates are presented in Table 1. The grass stand mainly consisting of peren-nial ryegrass (Lolium perenne) and meadow fescue (Fes-tuca pratensis) was harvested as f irst cut at mid vegetative stage (inflorescence emergence) using a disc mower at the research dairy farm of the University of Veterinary Medi-cine Vienna (Pottenstein, Austria). Thereby, a rather fiber-rich grass stand was intentionally selected to better study the fiber-cleaving effects of the novel silage additives. All si l ages were prepared with approximately 35% DM con centration and a compaction density of 212.5 kg DM/m’as recommended by the German Federal Working Group for Forage Preservation (Bundesarbeitskreis Futterkonser-vierung 2011). Thus, tap water was added to wheat straw before ensiling, whereas grass was wi l ted in the sun to adjust the DM concentration of plant material. Subsequently, the grass was chopped to 7 cm length before ensiling, while wheat straw was obtained from straw bales without mold Table 1 Chemical composi t ion of grass and wheat straw before ensil-ing Grass Wheat straw Dry matter (DM) concentration , g/kg 422 931 Ash, g/kg DM 101 68.0 Crude protein, g/kg DM 182 39.7 Ether extract, g/kg DM 20.1 11.6 aNDFom , g/kg DM 643 842 ADFom’, g/kg DM 286 473 ADL‘, g/kg DM 53.2 119 WSC,g/kg DM 167 66.6 Determined after wilting of grass and before water addi t ion to wheat straw Neutral detergent fiber assayed with a heat stable o-amylase and expressed exclusive of residual ash Acid detergent fiber expressed exclusive of residual ash 4Acid detergent l ignin Water-soluble carbohydrates or visible contamination and was already chopped to 7 cm particle length. Since straw is lacking water-soluble carbohy-drates (WSC), which constitute the metabolizable substrate for lactic acid bacteria and consequently are pivotal for a suf f icient acidi f ication in the silo (McDonald et al. 1991),18 g sucrose was added to each glass j ar, i .e., constituting 10% of DM ensiled. Regarding the applied treatments, both grass and straw were ensiled with either (a) 18 mL of heat-i nactivated fun-gal enzyme solution serving as a control (CON _AF),(b)18 mL of freshly prepared anaerobic fungal enzyme solu-tion (AF), (c) 18 mL of heat-inactivated mixed ruminal fluid serving as a control (CON _RF), or (d) 18 mL of freshly col-lected mixed ruminal fluid (RF). Therefore, the lyophilized powder, which contained the anaerobic fungal enzymes,was dissolved in dist i lled water and directly applied to the respective silages. The disti l led water was added in such a quantity that the same volume of supernatant was again obtained as before freeze-drying of fungal culture superna-tant. For the control treatments, both mixed ruminal fluid as well as dissolved AF supernatant were heat-inactivated by placing both into an oven at 103 °C for 4 h and subsequent cooling down to approximately 25℃ before being used as silage additives. The pH of the fresh and heat-i nactivated fungal enzyme solution as well as the fresh and heat-inac-tivated mixed ruminal f luid was 8.84, 8.37, 6.89, and 6.74,respectively. Consequently, eight different silage treatments were produced, which are referred to as G _CON _AF, G _CON _RF,S _CON _AF,S _CON _RF, G _AF, S _AF,G _RF,and S _RF. Each silage treatment was prepared in t riplicate and subsequently stored at 20±1.3 ℃ for 90 days. All silos were weighed on day 1 and day 90 to calculate the DM loss.Additionally, fresh samples of grass and wheat straw were collected directly before ensi l ing and kept at-20°C until further analysis. Analysis of silage composition 5.1.2). Regarding fiber fractions, propor t ions of neutral detergent f iber assayed with a heat stable a-amylase and expressed exclusive of residual ash (aNDFom), acid deter-gent f iber expressed exclusive of residua l ash (ADFom),and acid detergent lignin (ADL) was determined in a Fibretherm FT12 (Gerhardt GmbH & Co. KG, Konigswinter, Germany)according with methods 6.5.1, 6.5.2, and 6.5.3, respectively.The WSC concentration was analyzed in accordance with method 7.1.1. Analysis of silage fermentation pattern Directly after silo opening, cold-water extracts of all silages were prepared by mixing 50 g of silage sample with 100 mL of distilled water and placing it for 16 h in the fridge at 4°℃.Subsequently, the complete content was f i l tered through three layers of gauze (Wilhelm Weisweiler GmbH & Co.KG, Miinster, Germany) and pH was immediately deter-mined potentiometrically (S40-K SevenMultiTM pH meter,Mett l er Toledo, Vienna, Austria) in the liquid before being stored in aliquots at- 20 C until further analyses. The concentrations of volatile fatty acids acetate, pro pionate, butyrate, and ethanol were determined on a gas chromatography device (GC Model 8060 MS 172 DPFC,No.: 950713, Fisons, Rodano, Italy), equipped with a f lame-ionization detector and a 30-m×0.53-mm ID×0.53-um df capillary column (Trace TR Wax, Thermo Fisher Sci-entific, Vienna, Austria). The detailed protocol , including sample preparation, can be obtained from Hartinger et al.(2022). The ammonia concentration was analyzed using the Berthelot reaction (Hinds and Lowe 1980) and lactic acid concentration was determined by high-performance l iquid chromatography (UltiMate 3000 HPLC system, Thermo Fisher Scientific, Vienna, Austria) according to WeiB and Kaiser (1995). Determination of in situ degradability The effective ruminal degradability of DM, aNDFom, and ADFom of straw and grass silages as well as fresh straw and grass was determined applying the in situ nylon bag technique with a reduced number of incubation time points (Olaisen et al. 2003). Three runs were performed using the rumen-cannulated dry Holstein cow from which t he mixed ruminal f luid has been obtained. Beginning 14 days prior to the first nylon bag incubation, the cow was continuously fed a basal diet of grass silage and corn silage ad l ibitum (65:35, DM basis), which was provided in the morning and afternoon. Likewise , the cow was additionally fed 1 kg of concentrate mixture per day (i.e., ground corn, barley, and wheat in a ratio of 0.5:0.25:0.25) that was also provided in two equal portions with the basal diet. For the in situ incubations, each feedstuff, i.e., silages and fresh substrates, was dr i ed at 60 °℃ for 48 h and ground through a 3-mm screen in an ultra-centrifugal mill (ZM 200, Retsch, Haan, Germany). Subsequently, 10 g DM was weighed into 10-cm×20-cm polyester bags with a pore size of 50±10 um (R1020, ANKOM Technology, Macedon,NY,USA). Directly before being i ncubated in the rumen, dupli-cate samples of each feedstuff were placed in water (39°℃)for approximately 10 min with gentle movement of bags.Afterwards, all bags were inserted into the ventral sac of the rumen for 4 h, 12 h, 24 h, and 72 h. At the end of each incubation period, bags were removed and put in ice water to immediately stop microbial activity. Subsequently, all bags were washed on a cold rinse cycle in a washing machine for 30 min, dried at 60 ℃ for 48 h and analyzed for DM,aNDFom and ADFom as outlined before. Additionally, each run included“0 h bags”per feedstuff, which were directly removed af t er pre-soaking in water, i .e., not incubated into the rumen, and subsequently t reated similar as described for the other bags. The calculations of ruminal degradabil i ty of DM,aND-Fom, and ADFom were performed using the equation of McDonald (1981): where a constitutes the fraction that disappears from the bag i mmediately, b consti t utes the insoluble but potentially rumen-degradable fraction, c is the constant rate of disap-pearance of fraction b , t is the incubation period, and L rep-resents the lag phase. These non-linear parameters, i.e., a ,b, c, and L, were estimated using an iterative least squares procedure in SAS version 9.4 (SAS Institute Inc., Cary, NC,USA). The effective degradability of DM, aNDFom, and ADFom was estimated assuming a ruminal passage rate of 4%/h, as recommended for forages (Offner et al . 2003) using the equation: Effective ruminal degradability =a+b×c/(c+0.04) Statistical analysis Statistical analysis of silage composition and fermentation pattern variables was performed with the GLM procedure of SAS version 9.4 (SAS Institute Inc., Cary, NC, USA) using the following model: where u is the mean, p; is the main effec t of ensiled plant material , t; is the main effect of treatment, (p×t); is the two-way i nteraction between t he main effects, and e,; i s the residual error. Di f ferences between least square means were analyzed by Tukey-Kramer post hoc test. For the in si t u degradabi l ity data, statistical analysis was performed with the MIXED procedure of SAS version 9.4 (SAS Institute Inc., Cary, NC, USA) using the following model: where u is the mean, p; is the fixed effect of ensiled plant material, t; is the fixed effect of treatment, (p×t), i s the two-way interaction between the main effects, r is the random effect of run, and eijk is the residual error. Differ-ences between least square means were again analyzed by Tukey-Kramer post hoc test . The significance level was set at a=0.05, and a trend was declared at 0.050.05).Treating silages with AF supernatant reduced the pH by around 0.3 units (P=0.04). Besides, pH in grass silages was on a higher level than in straw silages (P<0.01), i .e.,4.72 vs. 3.87, respectively. The straw silages treated with AF supernatant had less acetic acid compared to its control,whereas acetic acid concentrations in grass silages were sim-ilar (P<0.01). Additionally, a substrate effect was observed (P<0.01), revealing that acetic acid was lower abundant in grass silages than in straw silages. Similarly, propionic acid concentration was also lower in grass silages than in straw s i lages (P=0.03). Regarding ethanol, a substrate effect was observed with higher concentrations in straw than in grass silages (P=0.04), whereas AF supernatant or its interac-tion with substrate had no impact. The ammonia proportion was generally i ncreased by the addition of AF supernatant (P<0.01) with higher values in S _AF than S _CON _AF (P=0.03). Besides, concentrations of ammonia were lower in grass silages than i n straw silages (P<0.01). No effects of AF supernatant treatment, substrate or their interaction were found for concentrations of ADL, WSC, lactic acid,and butyric acid (each P>0.05). Both the nutritional composition and fermentation pattern of grass and straw silages t reated with mixed ruminal fluid are presented in Table 3. Regarding the nutritional com-position, mixed ruminal fluid reduced the ADL concentra-tion compared to control silages, i.e., 64.4 vs. 118 g/kg DM (P<0.01), and tended to increase the ADFom concentration in silages (P=0.08). Moreover, the substrate affected several nutritional composition variables with higher concentrations of DM, ash, crude protein, and ether extract in grass silages compared to straw silages (each P<0.01), whereas al l f iber Table 3 Effect of mixed ruminal fluid on nutr i tional compos i tion, and fermentation pattern of silages Grass silage Straw silage SEM P-values CON_RF RF2 CON_RF RF Treatment Substrate Interaction DM'concentration, g/kg 409 386 342 348 0.80 0.31 <0.01 0.13 Ash, g/kg DM 105 111 68.1 63.6 2.84 0.78 <0.01 0.11 Crude protein, g/kg DM 187 190 36.1 45.4 3.56 0.14 <0.01 0.35 Ether extract, g/kg DM 26.7 28.7 13.0 10.8 1.38 0.94 <0.01 0.18 aNDFom", g/kg DM 553 576 832 842 8.78 0.10 <0.01 0.50 ADFom', g/kg DM 315 326 500 512 5.70 0.08 <0.01 0.94 ADL°, g/kg DM 93.1 47.3 142 81.4 6.70 <0.01 <0.01 0.30 WSC,g/kg DM 20.6 28.3 52.4 19.6 18.1 0.49 0.52 0.29 pH 4.60aB 4.29 4.19A 4.25 0.05 0.04 <0.01 0.01 Lactic acid, g/kg DM 22.7 84.87 8.85 46.0 11.7 0.01 0.07 0.30 Acetic acid, g/kg DM 16.2 29.6* 21.7 20.0 2.91 0.09 0.48 0.05 Butyric acid, g/kg DM 0.00 0.00 8.69 4.93 0.41 0.65 0.11 0.59 Propionic acid, g/kg DM 0.17 0.97B 1.27 4.13aA 0.35 <0.01 <0.01 0.03 Ethanol, g/kg DM 2.55B 4.60 8.44A 5.83 0.76 0.71 0.01 0.03 Ammonia, g/kg N 47.5 53.0 77.7b 104.8 4.59 0.01 <0.01 0.04 In each row, superscript capitalized letters indicate difference (P<0.05) between substrates within each treatment , i.e., CON _AF and AF, and superscript lowercase letters indicate di f ference (P <0.05) between t reatments wi t hin each substrate, i.e., straw and grass Silage prepared with 18 mL of heat-inactivated mixed ruminal fluid Silage prepared with 18 mL of freshly col l ected mixed ruminal fluid Dry matter “Neutral detergent f iber assayed with a heat stable o-amylase and expressed exclusive of residual ash Acid detergen t f i ber expressed exclusive of residual ash 6Acid detergent lignin Water-soluble carbohydrates fractions were higher in straw silages than in grass silages (each P<0.01). Regarding the silage fermentation pattern,mixed ruminal fluid lowered the pH in grass silages, whereas no differences were present within straw silages, but pH of S _CON _RF was lower than of G _CON _RF(P=0.01).Silage pH was also affected by main effects of mixed rumi-nal fluid treatment and substrate with higher pH in control silages (P=0.04) and grass si l ages (P<0.01), respectively.Similar to pH results , the acetic acid concentration in mixed ruminal f luid-treated grass silages was higher than in con-trol grass silages but not differing between straw silages (P=0.05). Contrastingly, the ammonia concentration was higher i n S _RF compared to S _CON _RF with similar values in grass silages (P=0.04). Besides, main effects of mixed ruminal fluid treatment and substrate revealed generally lower ammonia proportions in control silages (P=0.01)and grass silages (P <0.01), respectively. The addition of mixed ruminal fluid increased the lactic acid concentra-tion in both grass and straw silages (P=0.01). Moreover,grass silages tended to have more lactic acid compared to straw s i lages (P=0.07). The propionic acid concentration was higher i n mixed ruminal fluid-treated straw silages than control straw silages (P=0.03); plus within mixed ruminal fluid-treated silages, straw silage contained more propionic acid than grass silage (P=0.03). Besides, main effects of mixed ruminal fluid treatment and substrate were observed with higher propionic acid values for mixed ruminal fluid treatment than control (P<0.01) and straw silages than grass silages (P<0.01), respectively. The ethanol concentration was affected by the interaction of mixed ruminal f luid and substrate (P=0.03) with higher values in control straw silage than in control grass silage. Besides, a main substrate effect was observed for this variable and straw silages con-tained more ethanol than grass silages (P=0.01). Effects of mixed ruminal f luid treatment, substrate, or their interaction were not f ound for either WSC or butyric acid concentration (each P>0.05). In situ degradability The results of effective ruminal degradability regarding the use of AF supernatant as a silage additive are presented in Figs. 1 and 2; the respective data for constants a, b, and c and lag phase can be obtained from Table 4. The DM degradabil-ity was not different between G _CON _AF,G _AF, and fresh grass (Fig . 1A), whereas an interaction of substrate and AF supernatant treatment for DM degradabi l ity was observed (P<0.01) with higher values for S _CON _AF than S _AF and fresh straw (Fig. 2A ). Besides, also substrate affected DM degradability with higher values for grass than for straw (P<0.01). No main effect on DM degradability was observed for AF supernatant (P=0.55). An interaction of substrate and AF supernatant treatment was also present for A 60 50 20 10 0 F R E S H CON _A F A F Fig. 1 Ef f ective ruminal degradabi l ity (4%/h passage rate) of dry matter (A), neutral detergent fiber (B), and acid detergent fiber (C)of original fresh grass (FRESH) or grass silages prepared with heat-inactivated (CON _AF) or freshly prepared anaerobic fungi superna-tant (AF). Asterisk=P <0.05 in Tukey -Krame r post hoc test the aNDFom degradability (P<0.01) with lower values for S _AF than S _CON _AF and fresh straw (Fig. 2B), whereas for grass, aNDFom degradabil i ty was lower for G _CON _AF than G _AF and f resh grass (Fig. 1B). Moreover, main effects of substrate (P<0.01) and AF supernatant (P<0.01) were significant for ruminal aNDFom degradability. Regarding A 米 * 50 0 FRE SH CON _A F AF Fig.2 Effective ruminal degradability (4%/h passage rate) of dry matter (A), neutral detergent fiber (B), and acid detergent f i ber (C)of original f resh straw (FRESH) or straw silages prepared with heat-inactivated (CON _AF) or f reshly prepared anaerobic fungi superna-tant (AF). Asterisk=P <0.05 in Tukey-Kramer post hoc test the degradability of ADFom, interaction of substrate and AF supernatant was significant (P=0.01) with higher ADFom degradabi l ity for G _AF than for fresh grass with G _CON _AF being i ntermediate (Fig. 1C), whereas ADFom degradabil i ty of S _CON _AF was higher when compared to S _AF and fresh straw (Fig. 2C ). As found for aNDFom degradability, main effects of substrate (P<0.01) and AF supernatant (P=0.02) were also significant for ADFom degradability. The results of effective ruminal degradabil i ty regard-ing the use of mixed ruminal f luid as a silage inoculant are shown in Figs. 3 and 4; the respective data for constants a, b,and c and lag phase can be obtained from Table 5. Hereby,the effective ruminal DM degradabil i ty was influenced by substrate (P<0.01) with generally higher values for grass than for straw. The mixed ruminal fluid treatment (P=0.28)as well as the interaction of substrate and mixed ruminal fluid (P=0.45) showed no differences in DM degradability between incubated feedstuffs (Figs. 3A and 4A ). For aND-Fom degradabil i ty, a trend for the interaction of substrate and mixed ruminal f luid was found (P=0.08) with higher values for fresh grass compared to G _CON _RF and G _RF (Fig. 3B), but similar values for fresh straw, S _CON _RF,and S _RF (Fig. 4B ). Besides, substrate (P<0.01) and mixed rumina l fluid (P<0.01) affected the aNDFom degradabil-ity. As found for DM degradabi l ity, ADFom degradability was i nfluenced by substrate (P<0.01) with generally higher values for grass than for straw, whereas neither the mixed ruminal fluid treatment (P=0.41) nor its interaction with substrate (P=0.45) impacted the ADF degradability of incu-bated feedstuffs (Figs. 3C and 4C ). Discussion The present study investigated the impact of AF supernatant with active fungal enzymes on composition and fermenta-tion quality of grass and straw silages, i .e., a typical and a recalci t rant f iber-rich forage, and determined thei r ruminal DM and fiber degradability. We hypothesized a stronger lactic acid fermentation dur i ng ensiling as well as a higher ruminal fiber degradability due to enhanced cleavage of structural carbohydrates by AF supernatant in the silo. Regarding silage quality, the addition of AF superna-tant lowered the pH in all silages compared to respective controls, and in case of grass silage, the treatment reduced the silage pH to 4.54 and thus below the DM-dependent threshold for stable conservation (Muck 1988). Although the lactic acid level was not significantly affected, the numerical increase of 20.5 g/kg DM i n AF supernatant-treated grass silages seemed to - at least partly - explain this observed pH decline. Likewise, the AF supernatant decreased the acetic acid concentration i n straw silages without affecting the lactic acid concentration, indicating a shift towards a homolactic-dominated fermentation (Borreani et al . 2018).Consequently, these beneficial i nfluences on silage fermenta-tion characterist i cs revealed an improved silage quality and forage preservation with AF supernatant, which was also reflected by the trend for reduced DM losses in both grass fungi supernatant (S _AF) G_FRESH G_CON_AF G_AF S FRESH S_CON_AF S_AF Dry matter a 43.64 40.31 42.38 17.52 21.50 20.42 b2 43.46 48.63 48.10 57.67 58.34 54.45 c,%/h 0.06 0.05 0.06 0.03 0.03 0.03 Lag time 2.90 2.59 2.14 4.90 4.70 4.40 Neutral detergent fiber assayed with a heat stable o-amylase and expressed exclusive of residual ash C 37.59 23.28 30.32 10.59 11.64 9.79 b 51.16 63.96 58.20 71.16 67.59 69.60 c,%/h 0.05 0.04 0.05 0.03 0.03 0.02 Lag time 3.87 3.82 4.43 4.90 4.90 4.80 Acid detergent fiber expressed exclusive of residual ash C 16.47 20.15 23.12 7.50 16.83 13.00 b 81.41 66.00 62.74 62.16 61.16 52.75 c,%/h 0.03 0.04 0.05 0.02 0.03 0.02 Lag time 5.26 4.30 3.60 4.50 4.90 4.40 Fraction that disappears f rom t he bag immediately Insoluble but potent i ally rumen-degradable fraction 'Constant degradation rate of fraction b and straw silages. Therefore, our hypothesis of a stronger lactic acid fermentation may not be confirmed, but the overal l improved silage quality in response to AF superna-tant i ndeed suggest a beneficial effect of AF supernatant in silages. Interestingly, the addition of AF supernatant resulted ir an increased ammonia concentration in straw silages, sug-gesting higher protein degradation in the silo. I t is conceiva-ble that the AF supernatant as well i ncluded proteases as the applied AF supernatant was obtained from species of Neo-callimastix, a genus whose members partly show proteolytic activity, as well (Har t inger et al. 2018). Such AF-induced proteolysis is believed to be associated with the degrada-tion of structural proteins to sufficiently decompose fibrous plant structures, and i t may further modify the activities of other f ungus-derived CAZymes (Wal l ace and Jobl i n 1985).Therefore, a certain proportion of proteolytic activity i n AF cultures may be i nevitable, but in terms of f iber degrada-tion suppor t ive. However, i t has to be noted that ammonia proportion stil l amounted for less t han 10% of total nitrogen in AF supernatant-treated straw silages, which is deemed as a suf f icient t rue protein conservation (Kung et al. 2018).Similarly, no difference in ammonia concentrations between control and AF supernatant-t reated silages was observed for grass, meaning our positive assessment of AF supernatant for silage quality holds t rue. The proximate nutrients of the s i lages were mainly affected by the ensiled substrate, i.e., grass or straw,with expectedly higher concentrations of ash, crude pro-tein and ether extract , but lower concentrations of fiber fractions in grass than straw, as i t has also been found in the fresh substrates. The overall aNDFom concentrations were decreased after ensiling, which may be related to the acidic hydrolysis of hemicelluloses in the silo (Dewar et al.1963). The treatment with AF supernatant, however, showed no influence on the fiber fraction concentrations, meaning that a potential fiber tackling effect of AF enzymes was not reflected in silage composition. This is in contrast to prior studies inoculating rice straw or whole-plant corn with vari-ous viable AF species before ensi l ing, which observed less NDF and ADF concentrations compared to controls (Lee et al. 2015; Wang et al. 2019). The ruminal degradabi l ity of f iber fractions, however,was i ndeed affected by this treatment. Accordingly, our most i mportant finding here was t he improved fiber degra-dabi l ity of grass silages when ensiled with AF supernatant:The aNDFom degradability of AF supernatant-treated grass si l ages was higher t han of control grass silages and similar to fresh gras s , while ruminal ADFom degradabi l ity, i.e.,ruminal l i gnocellulose degradation, was even higher for AF supernatant-treated grass silages when compared to fresh grass. Thus, our hypothesis was confirmed and it can be assumed that the enzymes present in the AF supernatant pre-cleaved l ignocellulosic complexes in t he silos, thus allow-ing a higher fiber degradability, especially cellulose, and eventually higher energy exploitation from grass silage in the rumen. As outlined in our companion paper (Hartinger and Zebeli 2021), AF comprise a large enzymatic spectrum,and apart from p-glucosidase and endoxylanase, which have been considered i n our study, more enzymes or cellulosomes have likely been active in the applied AF supernatant. For instance, using transcriptomics and proteomics, Wang et al . A FRES H CO N _RF R F Fig.3 Effective ruminal degradability (4%/h passage rate) of dry matter (A), neutral detergent f iber (B), and acid detergent f iber (C) of fresh grass (FRESH) or grass si l ages prepared with heat-inac t ivated (CON _RF) or freshly prepared mixed ruminal fluid (RF). Aster-isk=P<0.05 in Tukey-Kramer post hoc test (2011) identified several novel f iber-degrading enzymes in Neocallimastix patriciarum W5. Thus, similar omics-based approaches can help to characterize the AF enzymes and consequently also to understand the modes of action in the silo. In case of straw silages, the beneficial effects of AF supernatant seen for silage quality could not be transferred to ruminal degradation. Surprisingly, the control straw silages, i.e., prepared with heat-inactivated AF supernatant, A 50 0 FRES H CO N _RF R F Table 5 Constants of ruminal degradation for fresh grass (G _FRESH), grass silage treated with heat-inactivated mixed ruminal f luid (G _CON _ RF), grass silage t reated wi t h freshly collected mixed ruminal f l uid (G _RF), fresh straw (S _FRESH), straw silage t reated with heat-inactivated mixed ruminal f luid (S _CON _RF),and straw silage treated with freshly collected mixed ruminal fluid (S _RF) G_FRESH G_CON_RF G_RF S_FRESH S_CON_RF S_RF Dry matter a 43.64 42.52 41.00 17.52 15.76 17.04 b2 43.46 45.70 49.47 57.67 55.91 50.59 c,%/h 0.06 0.06 0.05 0.03 0.02 0.04 Lag time 2.90 3.48 3.50 4.90 4.90 3.50 Neutral detergent fiber assayed with a heat stable o-amylase and expressed exclusive of residual ash C 37.59 22.55 19.50 10.59 6.71 10.96 b 51.16 66.61 69.28 71.16 76.03 62.14 c,%/h 0.05 0.04 0.05 0.03 0.02 0.03 Lag time 3.87 4.90 4.00 4.90 4.90 4.50 Acid detergent fiber expressed exclusive of residual ash C 16.47 21.68 20.08 7.50 6.94 11.17 b 81.41 68.66 65.38 62.16 59.91 48.97 c,%/h 0.03 0.04 0.05 0.02 0.01 0.03 Lag time 5.26 4.90 3.48 4.50 4.90 3.80 Fraction that disappears from the bag immediately Insoluble but potent i ally rumen-degradable fraction 'Constant degradation rate of fraction b et al. (2015), who observed higher ruminal fiber degrada-bility when ensi l ing rice straw with viable AF cultures. In contrast, wheat straw was ensi l ed in the present study, and apart f rom a di f ference between the application of viable AF cultures or their supernatant, a straw type-specific influence is possible and may be taken into account. Focusing only on the application of AF supernatant as a silage addi t ive, our results clearly showed a substrate-spe-cific influence on the efficacy of AF supernatant in silages,i.e., grass vs. straw. The AF supernatant i mproved the qual-ity of both grass and straw silages, but beneficial effects on ruminal f iber degradability were only present for grass silages. Thus, our hypothesis of a higher ruminal degrada-bility of forages ensiled with AF supernatant was confirmed for grass silages only. Studies investigating the impact of AF supernatant in other common forages, such as corn and alfalfa, can provide further information on substrate-speci f icities and therefore better define the potential areas of application. Likewise, refinements in the culturing and preparation of AF or their supernatant are required to further increase the efficacy of this novel silage additive. Hereby,co-culturing of AF and methanogens may consti t ute an option to additionally increase the f ibrolytic enzyme yield as suggested by the observed upregulated transcription of CAZymes in such co-cultures (Swift et al. 2019). Likewise,as previous data i ndicate di f ferences in AF species regarding their effectiveness as silage inoculants (Wang et al. 2019;Lee et al. 2015), research on applying supernatants from further AF species or genera seems rationale - although it may be noted that the present AF supernatant was obtained from a culture of three Neocallimastix species, a genus that is considered to express highest enzyme activities among AF cultures (Dagar et al . 2018), par t icularly after sequential sub-culturing (Ekinci et al, 2006). In regard to esterase activity,being mainly responsible for the breakup of l ignocellulosic complexes, a collection of AF supernatant after 1 or 2 days of cultivation may enhance its fiber cleaving impact as a silage inoculant, especially in straw silages, since esterase activities were observed to be highest during early fungal growth, whereas cellulase and xylanase peak at a later time point of incubation (Dagar et al.2018). As a second part, our study further assessed the use of mixed ruminal fluid as a silage additive in grass and straw silages, which may inoculate the silo with f i brolyt i c microbes and their enzymes that in consequence tackle f iber components during silo storage. Provided a similar efficacy as when using AF supernatant for ensi l ing, the approach of directly applying mixed rumina l f luid would mean a reduc-tion in complexity, time, and labor compared to the produc-tion of AF supernatant (Dollhofer et al. 2015). Consequently,we analyzed the same parameters as for AF supernatant-treated silages and expected an improved silage fermentation and subsequent ruminal fiber degradability in response to the microorganisms and enzymes deriving from the mixed ruminal fluid. The higher lactic acid concentrations in silages treated with mixed ruminal fluid indeed demonstrated an enhanced lactic acid fermentation. Likewise, addition of mixed rumi-nal fluid reduced the pH to 4.29 i n grass silages and there-fore indicated a stable forage conservation (Muck 1988).Compared to control, the acetic acid concentration was dou-bled in grass silages wi t h mixed ruminal f luid, i .e., 16.2 Vs. 29.6 g/kg DM, which suggested a higher activity of heterol-actic lactobacil l i or ruminal fluid-derived acetate producers.However , as shown by the lack of differences in DM losses,an acetic acid level of around 30 g/kg DM may not cause energy losses, but can actually be interpreted as beneficially in terms of aerobic stability due to yeast i nhibitory effects (Danner et al . 2003). Likewise, the overall low ethanol lev-els indicate a general reduced presence and suppression of epiphytic yeasts in al l si l ages (Kung et al . 2018), therefore supporting the assumed sufficient aerobic stability. The slightly higher ammonia concentrations observed in treated straw silages may have been caused by proteolytic microbes brought into the silo via mixed ruminal fluid (Hartinger et al. 2018; Puniya et al. 2015). Thus, the effects of mixed ruminal fluid on ammonia formation in straw silages were the same as those of AF supernatant and appear to have a similar rationale. In parts, the higher ammonia may have also originated from the mixed ruminal fluid i tself (Puniya et al. 2015) and vaporized during heat inactivation, thus co-explaining the lower ammonia level i n control straw silages Sti l l, the ammonia concentration of 10.5% of total nitrogen in mixed ruminal fluid-treated straw silages appears uncriti-cal in regards to true protein conservation (Kung et al. 2018)and, consequently, our hypothesis of an improved silage fermentation with the addition of mixed ruminal fluid was confirmed. Worth of notice is the apparently high l ignin reduction with mixed ruminal fluid inoculation in both grass and straw silages , which has not been observed for the treatment of silages with AF supernatant . Since the rumen microbiota is incapable or only minimally able to degrade lignin (Susmel and Stefanon 1993), this observation was indeed very sur-prising and lacks a direct explanation. It might be conjec-tured that the mixed ruminal f luid-induced ADL degradation led to a higher availability of fermentable sugars and there-fore would explain the increased lactic acid concentrations but absence of an effect of mixed ruminal fluid on WSC concentration - but this seems actually highly specula tive. In consequence, ruminal f iber degradabil i ty should be increased if lignin was truly degraded (Zabel and Morrell 2020). However, a beneficial effect of such a lignin reduc-tion was not reflected in ruminal degradabi l ity. In fact,the DM and ADFom degradability was not influenced by mixed ruminal fluid in grass and straw silages and,for aNDFom, ruminal degradability was even lower after ensiling grass with fresh or heat-inactivated mixed ruminal fluid when compared to fresh grass. Thus, our hypothesis of an improved f i ber degradation due to a pre-cleaving effect by ruminal fluid-derived microbes during ensi l ing could not be confirmed. However, as the DM degradability remained similar between fresh grass and grass silages,degradabil i ty of other nutrients must have compensated the reduction in aNDFom degradability, which may be investigated in future studies. Consequently, inoculation of grass and straw silages with mixed ruminal fluid may be suitable for improving silage quality, but for a f iber-cleaving activity in the silo, the targeted application of AF supernatant cannot be replaced by directly using mixed ruminal f luid . However, as demanded for AF supernatant,further research may help to optimize this approach, as well. For instance, a concentration of microorganisms and their enzymes via ultra f iltration of mixed ruminal fluid,like i t has been performed during processing of the AF supernatant, could increase the fiber cleaving potential during ensiling. Since the present straw silages were pre-pared of pure wheat straw, applying mixed ruminal f luid as a silage inoculant in mixed silages of straw and a second substrate, e.g., grass or WSC-rich by-products, may be worth of further investigation. In conclusion, thi s is the f irst study to investigate AF supernatant and mixed ruminal f luid as novel s i lage addi-tives for ensil i ng grass and wheat straw and we provide strong evidence that both candidates improve silage quality. The treatment with AF supernatant addi t ionally enhanced ruminal fiber degradability of grass silage,which should be associated with a f iber-cleaving fungal enzyme activity in the silo. Therefore, the application of AF supernatant in silages represents a promising strategy to support forage utilization by ruminants and should be pursued . Differences in ruminal degradabil i ty between wheat straw and grass silages suggest substrate-specific effects that need consideration in future research Acknowledgements The authors thank Anita Dockner, Suchitra Sharma, Sabine Leiner, and Manfred Hollmann from the Institute of Animal Nutr i tion and Functional Plant Compounds at the University of Veterinary Medicine, Vienna, for the assistance during the chemical analyses of samples, as well as Lenka Strosova from the I nstitute of Animal Physiology and Genetics, Czech Academy of Sciences, for t he AF supernatant preparation. Author contribution TH and QZ designed the research and acquired the funding. TH and KF conducted experiment and analyses . TH ana-lyzed the data and wrote the manuscript. KF and QZ reviewed and edited the manuscript . All authors read and approved the manuscript. Funding Open access funding provided by University of Veterinary Medicine Vienna . This study was funded by the Austrian Federal Min-istry of Agriculture, Regions and Tourism, Grant Number 101623. Data availability All data generated or analyzed during this study are included in this published ar t icle. Declarations Ethical statement All applicable international , national, and/or i nstitu-t i onal guidelines for t he care and use of animals were followed . Competing interests The authors declare no competing interests Open Access This article is licensed under a Creative Commons Attri-bution 4.0 Internationa l License, which permits use, sharing, adapta-tion, distribution and reproduction in any medium or format , as long as you give appropriate credi t to the original author(s) and the source,provide a link to the Creative Commons licence, and indicate i f changes were made. The images or other t hird party material in t his article are included in the article's Creative Commons licence , unless indicated otherwise in a credi t line to the material . If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds t he permitted use, you will need to obtain permission direct l y f rom the copyright holder . To view a copy of this l i cence , visi t http://creativecommons.org/licenses/by/4.0/. References Ambye-Jensen M, Johansen KS, Didion T, Kadar Z, Schmidt JE,Meyer AS (2013) Ensi l ing as biological pretreatment of grass (Festulol i um Hykor): the effect of composition, dry matter, and inocula on cel l ulose convertibi l ity. Biomass Bioenergy 58:303-312. https://doi .org/10.1016/j .biombioe.2013.08.015 Bidochka MJ , Tong KI , Khachatourians GG (1993) Partial puri f i-cation and characterization of two extracellular N-acetyl-D-glucosaminidases produced by the entomopathogenic fungus Beauveria bassiana. Can J Microbiol 39:40-45. http s://doi.org /10.1139/m93-006 Borreani G , Tabacco E, Schmidt RJ, Holmes BJ, Muck RE (2018)Silage review: factors affecting dry matter and quality losses in silages. J Dairy Sci 101:3952-3979.https://doi.org/10.3168/jds.2017-13837 Caldwel l DR, Bryant MP (1966) Medium without rumen fluid for nonselective enumeration and isolation of rumen bacteria .Appl Microbiol 14:794-801. ht tps ://doi.org /10.1128/am.14.5.794-801.1966 Dagar SS, Kumar S, Mudgil P, Puniya AK (2018) Comparative eval-uation of l ignocellulolytic ac t ivi t ies of f ilamentous cultures of monocentric and polycentric anaerobic fungi. Anaerobe 50:76-79.h t tps://doi.or g/10.1016/j.anaerobe.2018.02.004 Danner H, Holzer M, Mayrhuber E, Braun R (2003) Acetic acid increases stabi l ity of s i lage under aerobic conditions. Appl Environ Microbiol 69:562-567. https ://doi.org/10.1128/AEM.69.1.562-567.2003 Dewar WA, McDonald P, Whittenbury R (1963) The hydrolysis of grass hemicel l uloses during ensilage. J Sc i Food Agric 14:411-417. https://doi.org/10.1002/jsfa.2740140610 Dollhofer V, Podmirseg SM, Callaghan TM, Griffith GW, Fliegerova K (2015) Anaerobic fungi and their potential for biogas produc-tion. In: Giibitz GM, Bauer A, Bochmann G, Gronauer A, Weiss S (eds) Biogas science and technology. Springer, Cham, pp 41-61 Ekinci M, Ozkose E, Akyol I (2006) Effects of sequential sub-cul-turing on the survival and enzyme activity of Neocallimastix hurleyensis. Turk J Biol 30:157-162 Forzieri G, Feyen L, Rojas R, Florke M, Wimmer F, Bianchi A (2014) Ensemble projections of future streamflow droughts in Europe . Hydro l Earth Sys t Sci 18:85-108. https://doi.org/10.5194/hess-18-85-2014 Futterkonservierung B (2011) Praxishandbuch Futter- und Substrat-konservierung, 8th edn. DLG-Verl , Frankfur t am Main Gardes M, Bruns TD (1993) ITS primers with enhanced speci f icity for basidiomycetes-application to t he identification of mycor-rhizae and rusts. Mol Ecol 2:113-118. htt ps ://doi.org /10.1111/j.1365-294x .1993.tb00005.x Hagen LH, Brooke CG, Shaw CA, Norbeck AD, Piao H, Arntzen MO,Olson HM, Copeland A, Isern N, Shukla A, Roux S, Lombard V,Henrissat B, O'Malley MA, Grigoriev IV, Tr i nge SG, Mackie RI, Pasa-Tolic L, Pope PB, Hess M (2020) Proteome specialization of anaerobic fungi during ruminal degradation of recalcitrant plant f i ber. ISME J. https://doi.o rg/10.1038/s41396-020-00769-X Hai t jema CH, Gilmore SP, Henske JK, Solomon KV, de Groot R,Kuo A, Mondo SJ , Salamov AA, LaButti K, Zhao Z, Chiniquy J, Barry K, Brewer HM, Purvine SO, Wright AT, Hainaut M,Boxma B, van Alen T, Hackstein JHP, Henrissat B, Baker SE,Grigoriev IV,O'Mal l ey MA (2017) A parts list for fungal cel-lulosomes revealed by compara t ive genomics. Nat Microbiol 2:17087. ht tp s://doi.org/10.1038/nmicrobiol.2017.87 Haitjema CH, Solomon KV, Henske JK, Theodorou MK, O'Malley MA (2014) Anaerobic gut fungi: advances i n i solation, culture,and cellulolytic enzyme discovery for biofuel production. Bio-technol Bioeng 111:1471-1482. https://doi.org/10.1002/bit.25264 Hartinger T, Grabher L, Pacifico C, Angelmayr B, Faas J , Zebeli Q (2022) Short-term exposure to the mycotoxins zearalenone or fumonisins affects rumen fermentation and microbiota, and health variables in cat t le. Food Chem Toxicol 162:112900.https://doi.or g/10.1016/j.fct.2022.112900 Hartinger T, Gresner N, Sudekum K-H (2018) Does intra-ruminal nitrogen recycling waste va l uable resources? A review of major players and thei r manipulation. J Anim Sci Biotechnol 9:33.h tt ps://doi.or g/10.1186/s40104-018-0249-x Hartinger T , Zebel i Q (2021) The present role and new potentials of anaerobic fungi in ruminant nutr i tion. J Fungi (basel ) 7:200.h tt ps://doi.or g /10.3390/jof7030200 Hess M, Paul SS , Puniya AK, van der Giezen M, Shaw C, Edwards JE, Fliegerova K (2020) Anaerobic fungi: past , present , and future . Front Microbiol 11. https://doi.org/10.3389/fmicb.2020.584893 Hinds AA, Lowe LE (1980) Appl i cation of the Berthelot reaction to the determination of ammonium-N in soi l extracts and soil digests.Commun Soil Sc i Plant Anal 11:469-475. https://doi.org/10.1080/00103628009367054 Kung L, Shaver RD, Grant RJ, Schmidt RJ (2018) Silage review: inter-pretation of chemical, microbial, and organoleptic components of silages. J Dairy Sci 101:4020-4033. https://doi .org /10.3168/ids.2017-13909 Lauer J (2012) The effects of drought and poor corn pollination on corn. Field Crops 28:493-495 Lee SM, Guan LL, Eun J-S, Kim C-H, Lee SJ, Kim ET,Lee SS (2015)The effec t of anaerobic fungal inoculation on the fermentation characterist i cs of rice straw silages. J Appl Microbiol 118:565-573. https://doi.o rg /10.1111/j am .12724 Lever M (1977) Carbohydrate determination with 4-hydroxybenzoic acid hydrazide (PAHBAH): effect of bismuth on the reaction.Anal Biochem 81:21-27. https://doi.org /10.1016/0003-2697(77)90594-2 McDonald P (1981) A revised model for the estimation of protein deg-radability in the rumen. J Agric Sci 96:251-252. https://doi.org/10.1017/S0021859600032081 McDonald P, Henderson N, Heron S (1991) The biochemistry of silage,2nd edn. Chalcombe Publications, Marlow, Bucks, UK Muck RE (1988) Factors influencing silage quality and thei r i mplica-tions for management. J Dairy Sci 71:2992-3002. https://doi.org/10.3168/jds.S0022-0302(88)79897-5 Mura E, Edwards J, Kittelmann S, Kaerger K, Voigt K, Mrázek J,Moniello G, Fliegerova K (2019) Anaerobic fungal communities differ along the horse digestive tract. Fungal Biol 123:240-246.h tt ps://doi.or g/10.1016/j.funbio.2018.12.004 Offner A, Bach A, Sauvant D (2003) Quantitative review of in situ starch degradation i n the rumen. Anim Feed Sc i Techno l 106:81-93. https://doi.o rg /10.1016/S0377-8401(03)00038-5 Olaisen V, Mejdel l T, Volden H, Nesse N (2003) Simpli f ied in situ method for estimating ruminal dry matter and protein degradability of concentrates. J Anim Sci 81:520-528. http s://doi .org/10.2527/2003.812520x Puniya AK, Singh R, Kamra DN (ed) (2015) Rumen microbiology:from evolution to revolution. Springer India, New Delhi , India Susme l P, Stefanon B (1993) Aspects of l ignin degradation by rumen microorganisms. J Biotechnol 30:141-148. https://doi.org/10.1016/0168-1656(93)90035-L Swi f t CL, Brown J L, Seppala S, O'Mal l ey MA (2019) Co-cul t ivation of the anaerobic fungus Anaeromyces robustus with Methano-bacterium bryantii enhances transcription of carbohydrate active enzymes. J Ind Microbiol Biotechnol 46:1427-1433.https://doi.org/10.1007/s10295-019-02188-0 VDLUFA (2012) VDLUFA-Methodenbuch Bd. II I Die chemische Untersuchung von Futtermitteln, 3rd edn. VDLUFA-Verlag,Darmstadt , Germany Wallace R, Joblin KN (1985) Proteolytic activity of a rumen anaerobic fungus. FEMS Microbiol Let t 29:19-25. https://doi.org /10.1111/.1574-6968.1985.tb00828.x Wang D, Zhao C, Liu S, Zhang T, Yao J , Cao Y (2019) Effects of Piromyces sp . CN6 CGMCC 14449 on fermentation quality,nutrient composition and the in vitro degradation rate of whole crop maize silage. AMB Express 9:121. https://doi.org/10.1186/s13568-019-0846-X Wang T-Y, Chen H-L, Lu M-YJ, Chen Y-C, Sung H-M, Mao C-T, Cho H-Y, Ke H-M, Hwa T-Y , Ruan S-K, Hung K-Y, Chen C-K, LiJ-Y,Wu Y-C, Chen Y-H, Chou S-P, Tsai Y-W, Chu T-C, Shih C-CA,Li W-H, Shih M-C (2011) Functional characterization of cellulases identi f ied from the cow rumen fungus Neocallimastix patriciarum W5 by transcriptomic and secretomic analyses. Biotechnol Biofu-els 4:1-16. https://doi.or g/10.1186/1754-6834-4-24 WeiB K, Kaiser E (1995) Milchsaurebestimmung in Silageextrakten mit Hi l fe der HPLC. Wirtschaftseig Futter 41:69-80 WeiBbach F, Kuhla S (1995) Stoffverluste bei der Bestimmung des Trockenmassegehaltes von Silagen und Gri i nfutter - entstehende Fehler und Moglichenkeiten der Korrektur. Uebers Tierernaehr 23:189-214 White TJ, Bruns T , Lee S, Taylor J (1990) Ampli f ication and direct sequencing of f ungal ribosomal RNA genes for phylogenetics. In:Innis M, Gelfand D, Sninsky J , White T (eds) PCR protocols : a guide to methods and applications. Academic Press, United States,pp 315-322 Zabel RA, Morrell JJ (2020) Chemical changes in wood caused by decay fungi . In: Morrell JJ , Zabel RA (eds) Wood microbiology 2E: decay and its prevention. Academic Press, Cambridge, USA,pp 215-244 Zhao J , Dong Z, Li J , Chen L, Bai Y, Jia Y, Shao T (2018) Ensiling as pretreatment of rice straw: The effect of hemicellulase and Lac-tobacillus plantarum on hemicel l ulose degradation and cellulose conversion. Bioresour Technol 266:158-165. https://doi .org/10.1016/j.biortech.2018.06.058 Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in publ i shed maps and institutional affi l iations.
确定







还剩12页未读,是否继续阅读?

产品配置单







中国格哈特为您提供《青贮饲料中α淀粉酶处理过的扣灰中性洗涤纤维 (aNDFom), 扣灰酸性洗涤纤维 (ADFom), 酸洗木质素(ADL)以及蛋白质含量的检测》,该方案主要用于饲料中营养成分检测,参考标准--,《青贮饲料中α淀粉酶处理过的扣灰中性洗涤纤维 (aNDFom), 扣灰酸性洗涤纤维 (ADFom), 酸洗木质素(ADL)以及蛋白质含量的检测》用到的仪器有格哈特全自动型纤维分析仪FT12、格哈特凯氏消化系统KT8S、格哈特维克松废气实验室废物处理系统涤气VS、格哈特全自动快速索氏提取SOXTHERM、格哈特带自动进样器凯氏定氮仪VAP500C、格哈特快速纤维测定器FBS6、德国加液器MM















相关方案
更多
该厂商其他方案
更多