







欧洲梨(Pyrus communis L.)在低温下贮藏以延长采后寿命。不幸的是,长时间暴露在贮藏环境中很可能发生老化烫伤和内部破裂。1-MCP能控制成熟障碍。本研究的主要目的是研究1-MCP和CA气调对冷藏‘Cold Snap’和‘Swiss Bartlett’梨果实品质的影响,包括生理障碍和氧化应激代谢产物。用或不加1-MCP处理鲜采梨,然后在0℃冷藏空气或CA气调(18kpa或2.5kpa氧气和2kpa CO2)下贮藏至少167d
方案详情

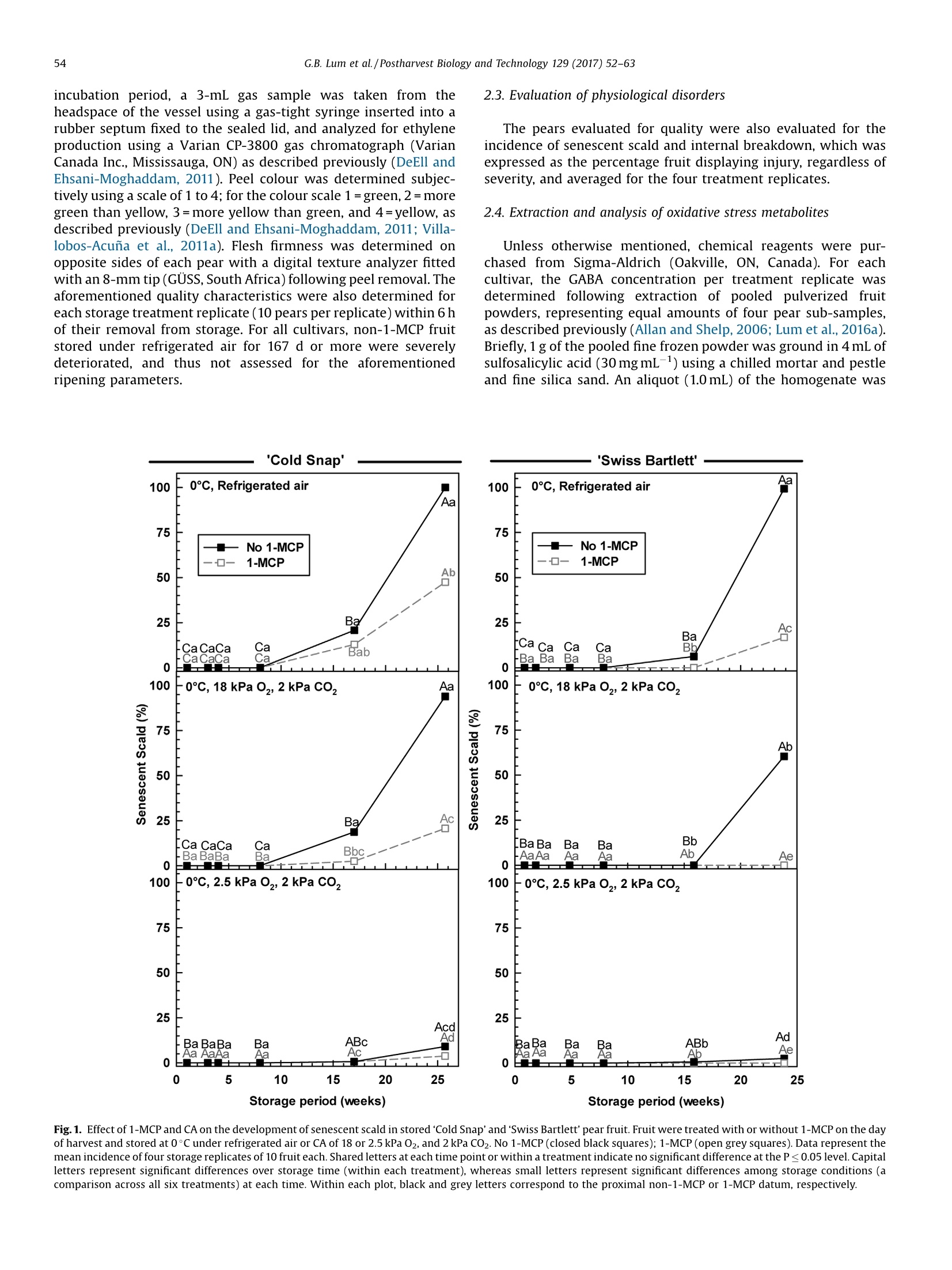
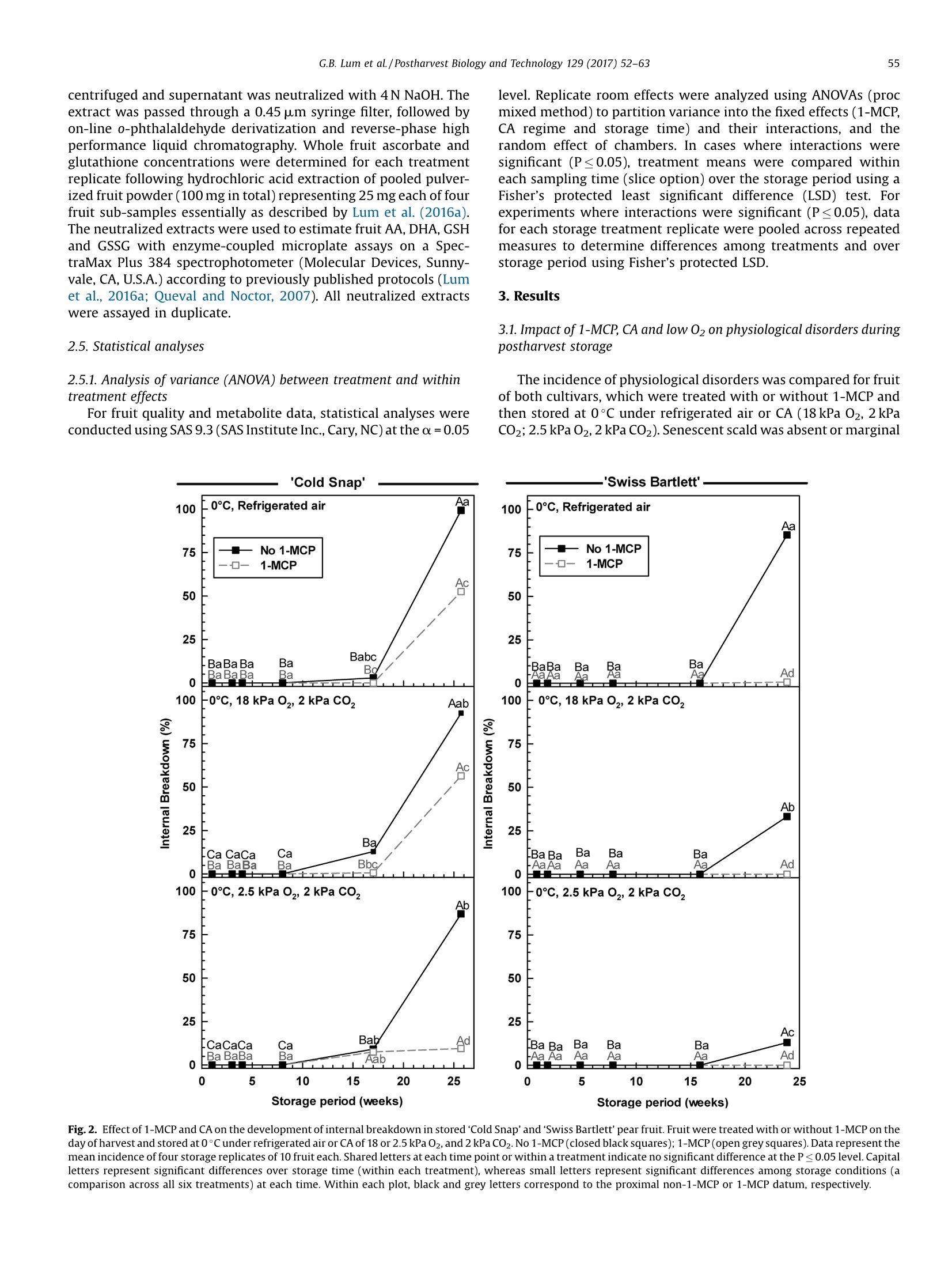
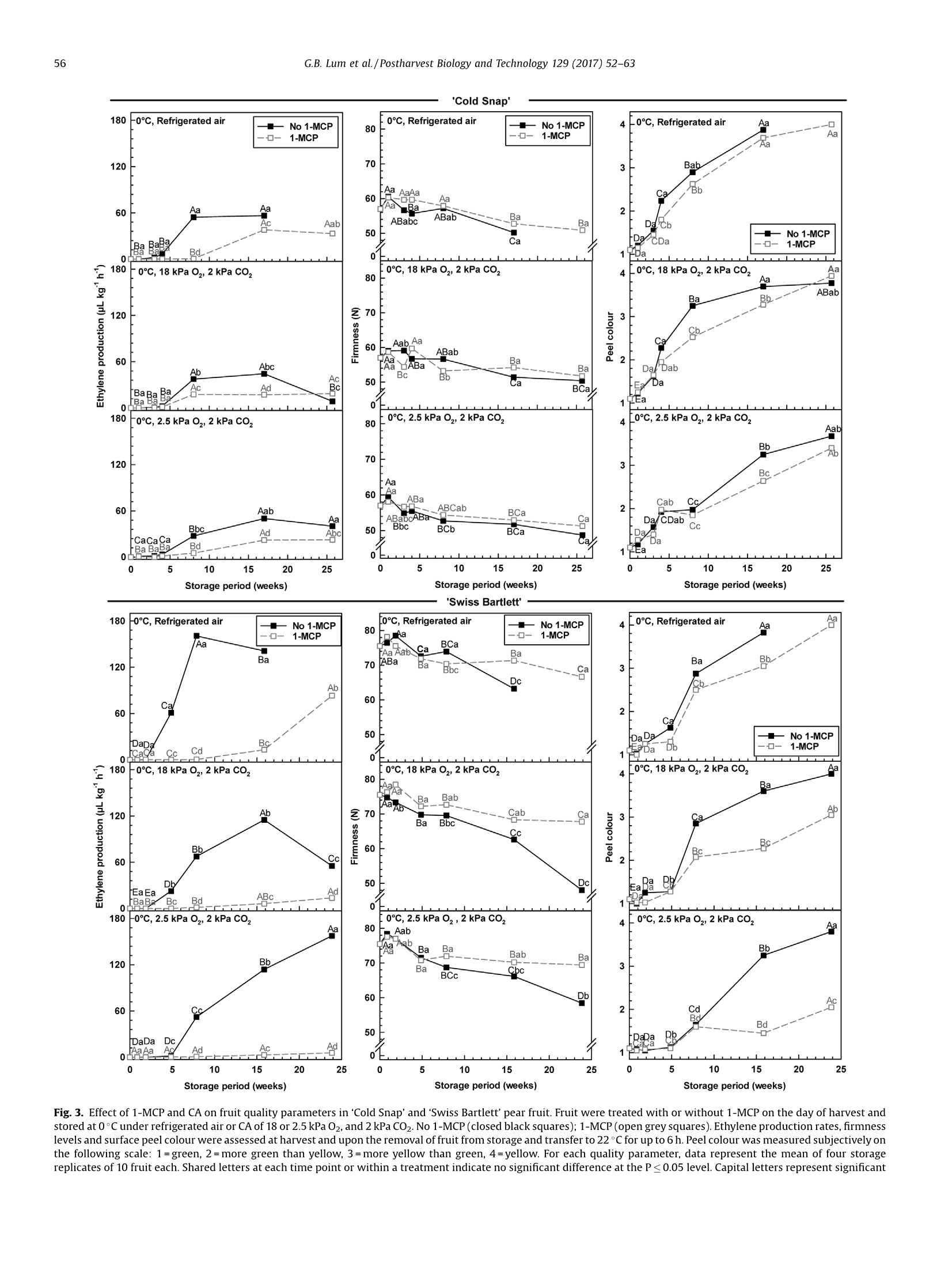
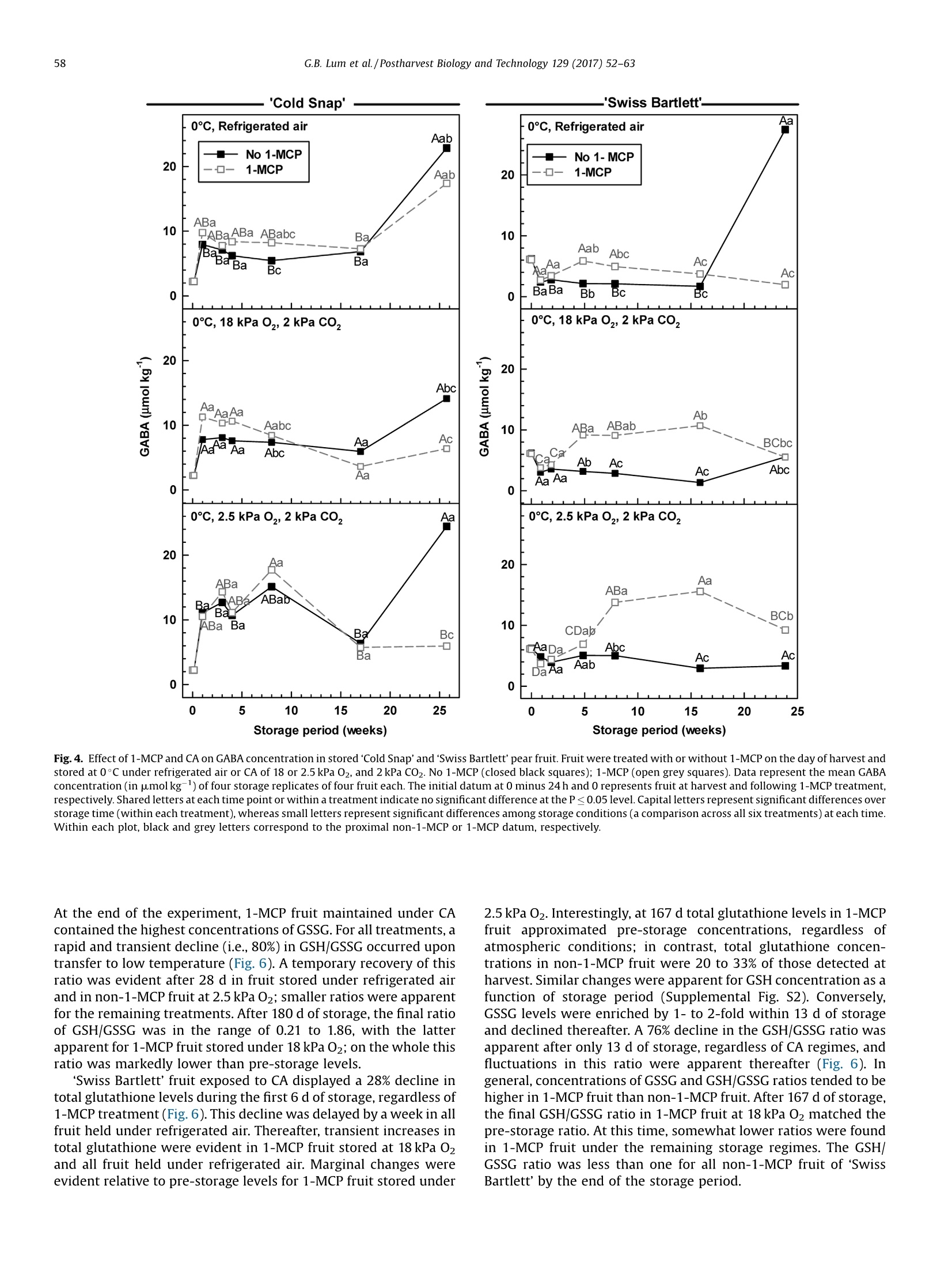
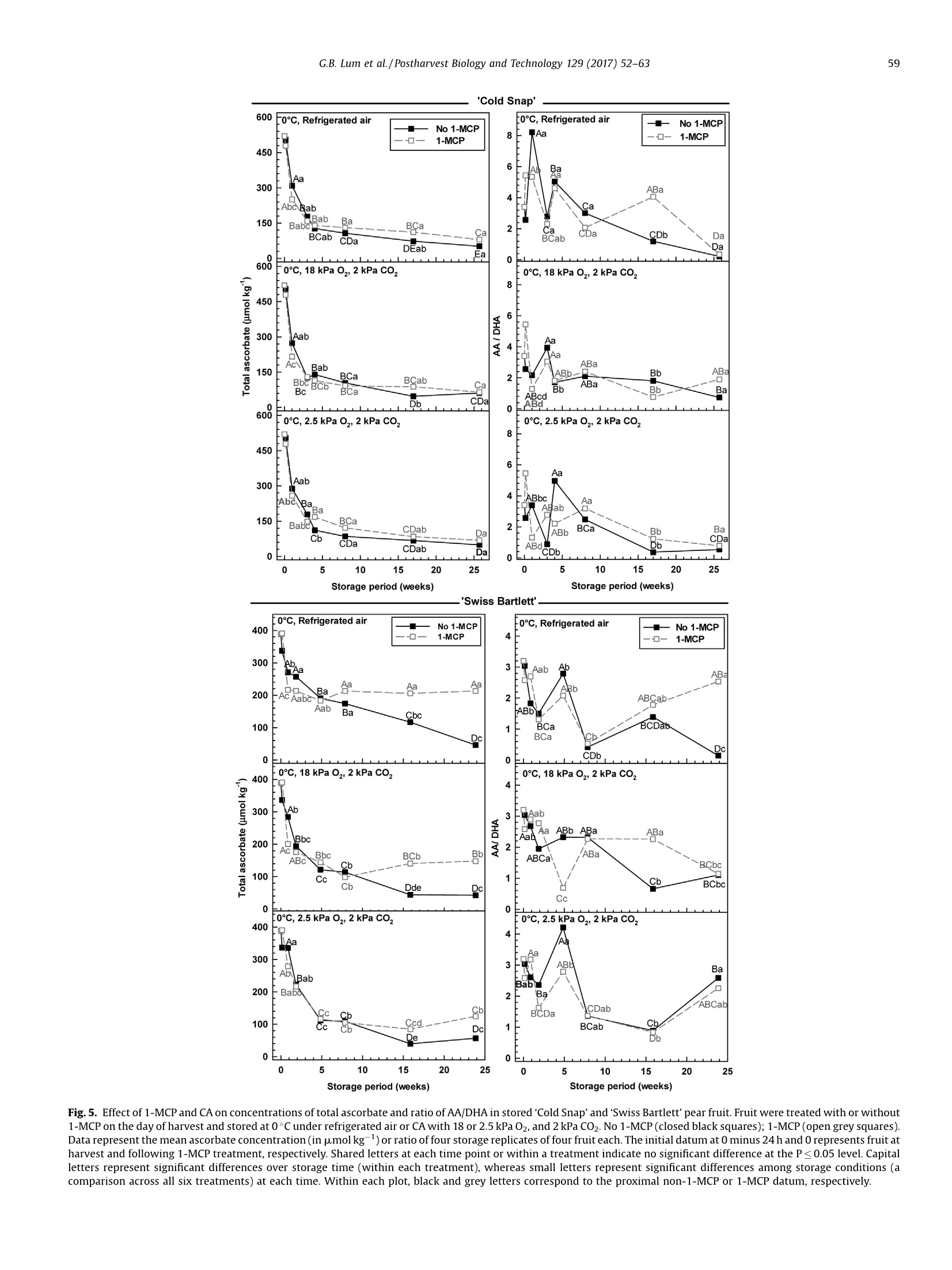
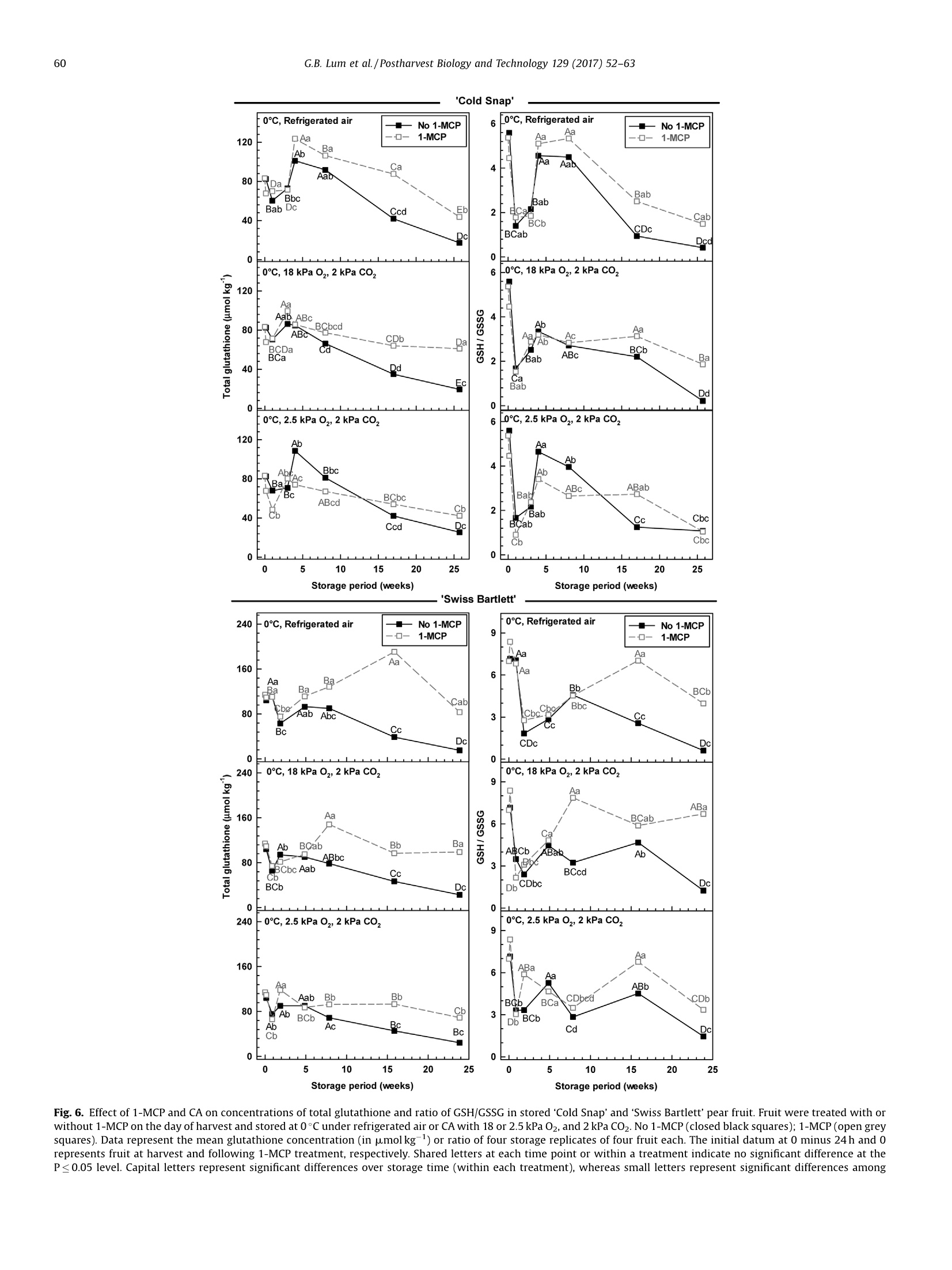
Postharvest Biology and Technology 129 (2017)52-63Contents lists available at ScienceDirect 53G.B. Lum et al./Postharvest Biology and Technology 129 (2017) 52-63 Postharvest Biology and Technology ELSEVIER journal homepage: www.elsevier.com/locate/postharvbio 1-Methylcylopropene and controlled atmosphere modulate oxidativestress metabolism and reduce senescence-related disorders in storedpear fruit CrossMark Geoffrey B. Lum, Jennifer R. DeEll, Gordon J. Hoover, Sanjeena Subedi .,Barry J. Shelp, Gale G. Bozzo ,* Department of Plant Agriculture, University of Guelph, 50 Stone Rd E., Guelph, ON, N1G 2W1, Canada BOntario Ministry of Agriculture Food and Rural Affairs, 1283 Blueline Rd. at Highway 3, Simcoe, ON, N3Y 4N5, Canada Department of Mathematics and Statistics, University of Guelph, 50 Stone Rd E., Guelph, ON, N1G 2W1, Canada European pears (Pyrus communis L.) are stored under low temperatures to extend postharvest life.Unfortunately,senescent scald and internal breakdown are likely to occur with prolonged exposure tostorage. Both disorders can be reduced by controlled atmosphere (CA) and/or the ripening inhibitor,1-methylcylcopropene (1-MCP). The principal aim of this study was to investigate the effect of 1-MCP andCA on fruit quality, including physiological disorders, and oxidative stress metabolites in stored ‘ColdSnap’and 'Swiss Bartlett' pears. Freshly harvested pears were treated with or without 1-MCP, and thenstored at O°C under refrigerated air or CA (18 kPa or 2.5 kPa 02, and 2 kPa CO2) for at least 167 d. 1-MCPand CA delayed and/or reduced the rates of ethylene production in stored fruit of both cultivars. 1-MCPand CA delayed fruit softening and peel yellowing in ‘Swiss Bartlett’ pears, but had negligible to slighteffects with ‘Cold Snap’. In both cultivars, high incidences of senescent scald and internal breakdownoccurred in non-1-MCP-treated pears during refrigerated air storage. For the most part these symptomswere reduced by CA and 1-MCP, resulting in minimal to negligible incidence in 1-MCP-treated pearsstored at 2.5 kPa 02.y-Aminobutyrate accumulated in stored pears, although 1-MCP and CA slightlyreduced the levels in'Cold Snap'fruit and 1-MCP increased levels in 'Swiss Bartlett' fruit. Ascorbate (totaland reduced) levels were rapidly depleted in ‘Cold Snap'fruit, regardless of treatment; these levels werebetter maintained in 1-MCP-treated ‘Swiss Bartlett’fruit than control fruit across all storageatmospheres. In both cultivars, glutathione (total and reduced) concentrations and redox statusfluctuated during storage,although these levels were generally higher in 1-MCP-treated fruit. Moreover,glutathione depletion occurred in advance of the development of senescence disorders in stored pearfruit. O 2017 Elsevier B.V. All rights reserved. 1. Introduction European pear (Pyrus communis L.) belongs to the Rosaceaefamily and is the fifth most cultivated tree fruit commodity inNorth America (FAOSTAT, 2016). The cultivar ‘Bartlett'represents75% of all pears that are grown in this region (Ingels et al., 2007). Amajor challenge for the production of ‘Bartlett' and its strain 'Swiss ( * Corresponding author. E-mail address: gbozzo @ uo gue lph.ca (G.G. Bozzo). ' Present address: Department of Mathematical Sciences, Binghamton University (State University o f New York), 4440 Vestal Parkway E ., Binghamton, N Y, 1 3902, USA. ) Bartlett'is their susceptibility to fire-blight in the orchard (Bonnandl van der Zweet, 2000; Hunter and Slingerland, 2008).Therefore, new cultivars with resistance to fire-blight have beendeveloped over the last decade. The fire-blight resistant pearcultivar ‘Cold Snap'(formerly ‘Harovin Sundown'; Harris, 2016)was developed by crossing ‘Bartlett'with US56112-146 (Hunteret al., 2009). To date, it is not known whether storability of pearsdiffers between fire-blight resistant and susceptible cultivars. Inorder to extend the market for freshly harvested pear fruit, mostEuropean cultivars are stored under low temperature (-1 to 10°C),including controlled atmosphere (CA) for periods of 3-10 months(Kader, 2007; Kupferman, 2003; Villalobos-Acuna and Mitcham,2008). A consequence of prolonged1:storagecanbethe development of internal breakdown and senescent scald in'Bartlett'and ‘Cold Snap'pear fruit (DeEll and Ehsani-Moghaddam,2011; Lum et al., 2016b). Symptoms of internal breakdown (alsoknown as internal browning and core breakdown) first appear asbrowning of the core tissue followed by cortical browning (Lumet al., 2016b; and references therein). Senescent scald is browningof the peel surface that can extend into the underlying flesh, and isetiologically distinct from superficial scald (Lum et al.,2016b; Lurieand Watkins, 2012). Superficial scald develops during the post-storage warming period and occurs as irregular brown patches ofthe peel that can become rough; it is associated with accumulationof the volatile a-farnesene and its subsequent oxidation (Lurie andWatkins, 2012). Interestingly, senescent scald is not associatedwith the accumulation of these metabolites in‘Abbe Fétel' pears(Vanoli et al., 2010). Both internal breakdown and senescent scaldcoincide with ethylene production in stored pears (Argenta et al.,2016; DeEll and Ehsani-Moghaddam, 2011; Ekman et al., 2004). Ethylene-mediated ripening and senescence of pear fruit areeffectively reduced by CA, specifically the use of low 02 partialpressures of 1-2.5 kPa and elevated CO2 partial pressures of 0.5-5 kPa (Drake, 1994; Kader, 2007; Kupferman,2003). Unfortunately,CA can promote core breakdown in Conference'and ‘Rocha'pears(Deuchande et al., 2016; Lum et al., 2016b). A complementarystrategy to CA is the pre-storage application of the ripeninginhibitor 1-methylcyclopropene(1-MCP)(Watkins, 2006,2015).1-MCP-treated ‘Bartlett’ pears are less susceptible to internalbreakdown and senescent scald during low temperature storagethan non-1-MCP-treated fruit (DeEll and Ehsani-Moghaddam,2011; Wang and Sugar, 2015). Pear fruit, including Cold Snap',arefree of these senescence disorders when 1-MCP treatment iscombined with CA; a drawback of these postharvest treatments isthe dramatic reduction or inhibition of ethylene-mediatedripening (Gago et al., 2015; Lum et al., 2016b; Ma and Chen,2003; Villalobos-Acuna and Mitcham, 2008). The biochemicalmechanisms associated with the development of senescent scaldand internal breakdown in Bartletttype pear fruit, and theircontrol by 1-MCP and CA, are not well understood. In stored pear fruit, a temporal reduction in respiration-derivedATP coincides with an increase in fermentation-related metabo-lites and NAD(P)H (Lum et al.,2016b). NADPH is used to generatereactive oxygen species and for the catabolism of y-aminobutyrate(GABA) (Foyer and Noctor, 2011; Lum et al., 2016b). Lowtemperature and hypoxia cause the accumulation of GABA inapple and pear fruits, which coincides with physiological disorders(Leisso et al., 2015; Lum et al., 2016a,2016b). Alternatively, GABAlevels are rapidly elevated in CA-treated plant tissues and this canoccur in the absence or prior to the appearance of visible injurysymptoms (Lum et al., 2016b; Zhou et al., 2016). In stored pome fruit, a shift in cellular NADPH redox statusculminates in a decline in the antioxidant capacity within the fruit,as this cofactor is used to power the ascorbate-glutathionerecycling pathway to generate reduced glutathione (GSH) andreduced ascorbate (AA) required for sequestering reactive oxygenspecies (Lum et al., 2016a, 2016b). Decreased ascorbate levels occurin advance of core breakdown and internal browning in pears,butin some cultivars this is evident in the absence of disorders(Deuchande et al., 2016; Lum et al.,2016b). Similarly, glutathionelevels and its redox balance tend to fluctuate in CA-stored pearfruit, and can contribute to the occurrence of physiologicaldisorders, including CA-related injury in ‘Honeycrisp’apples(Lum et al., 2016a,2016b). To date, no information exists onwhether shifts in ascorbate, GABA and glutathione are linked to thedevelopment of senescent scald and internal breakdown. In this study, we investigated the effect of 1-MCP and CA (18 kPaor 2.5 kPa O2 in the presence of 2 kPa CO2) versus refrigerated air onthe incidence of senescent scald and internal breakdown in pears of two genotypes, ‘Swiss Bartlett'and ‘Cold Snap', as a function ofstorage period at O°C. The near ambient O2 partial pressure of18 kPa was chosen as a control for the low O2 treatment and itscorresponding impact on disorder development and oxidativestress metabolism. The temporal development of senescencedisorders in these pear fruit was compared to changes in wholefruit concentrations of GABA, AA, dehydroascorbate (DHA), GSHand glutathione disulphide (GSSG). 2. Materials and methods 2.1. Plant material and postharvest treatments In 2012, pear fruit were harvested during the commercialharvest period. ‘Swiss Bartlett’ pears (Pyrus communis L.) wereharvested on August 22 from a commercial orchard near PortBurwell,ON, Canada, whereas ‘Cold Snap’ pears were harvested onSeptember 11 from a commercial orchard near Simcoe, ON,Canada. For each cultivar, the harvested material was randomlydivided into 24 boxes (~17 kg of fruit per box) for postharvesttreatments. Pears were transported to the storage research facility at theUniversity of Guelph (Guelph, ON, Canada) within 4h of theirharvest. For each cultivar, 12 boxes containing pears were equallydistributed into three separate air-tight 6 mil polybags, andexposed to 300 nLL-1-MCP for 24 h at the ambient temperatureof 23°C. The 1-MCP gas concentration used for each polybagtreatment is equivalent to the registered label rate for Smart-FreshM in Canada, and was calculated as described previously(Lum et al., 2016a). A similar quantity of boxes not exposed to 1-MCP remained at 23°C for 24 h. Following 1-MCP exposure, polybags were opened andventilated. A complete randomized block design was used forthe storage experiment to eliminate the possibility of room andchamber effects. Four separate temperature-controlled roomswere set to 0°C, each containing two lab-style CAchambers(0.5 m’volume; Storage Control Systems, Sparta, MI, United States). EachCA chamber contained one box each of ‘Cold Snap'pears treatedwith 1-MCP (1-MCP fruit) and one box of non-1-MCP fruit (non-1-MCP fruit). In addition, each CA chamber contained one box each of‘Swiss Bartlett' fruit treated with 1-MCP and one box ofnon-1-MCPfruit. All CA chambers were sealed with a plexiglass lid and flushedwith 98 kPa N2 until the target O2 concentration (18 kPa or 2.5 kPa02) was realized. Thereafter, CO2 was added until a partial pressureof 2 kPa was achieved;this CO2 concentration was chosen as anintermediate partial pressure to the range (0.5-5 kPa CO2) that istypically used for prolonged storage of pear fruit in otherjurisdictions (Kader, 2007). Also, one box of 1-MCP fruit and onebox of non-1-MCP fruit from each cultivar were stored within eachcold room without CA (refrigerated air). For each cultivar andtreatment, fruit were sampled periodically during the storageperiod. ‘Swiss Bartlett' fruit were sampled at 6,13, 34,55,111, and167 d, whereas ‘Cold Snap'fruit were sampled at 7, 21, 28, 56, 119and 180 d. For each sampling, 14 pears were chosen randomly forassessment of quality, including physiological disorders (10 pears)and metabolite analysis (four pears) as described below. Fruitsampled for metabolite analysis were immediately frozen in liquidN2 and stored at-80°c. 2.2. Evaluation of fruit quality Immediately after harvest, fruit maturity was assessed on eachof three biological replicates (replicate represented fruit from oneof three distinct locations within the orchard) of 10 pears. For eachreplicate, three representative pears were transferred to a 9Lplastic vessel and sealed air-tight for 60 min. At the end of the incubation period, a 3-mL gas sample was taken from theheadspace of the vessel using a gas-tight syringe inserted into arubber septum fixed to the sealed lid, and analyzed for ethyleneproduction using a Varian CP-3800 gas chromatograph (VarianCanada Inc., Mississauga, ON) as eFdescribed previously (DeEll andEhsani-Moghaddam, 2011). Peel colour was determined subjec-tively using a scale of 1 to 4; for the colour scale 1=green, 2=moregreen than yellow, 3=more yellow than green, and 4=yellow, asdescribed previously (DeEll and Ehsani-Moghaddam, 2011; Villa-lobos-Acuna et al., 2011a). Flesh firmness was determined onopposite sides of each pear with a digital texture analyzer fittedwith an 8-mm tip (GUSS, South Africa) following peel removal. Theaforementioned quality characteristics were also determined foreach storage treatment replicate (10 pears per replicate) within 6 hof their removal from storage. For all cultivars, non-1-MCP fruitstored under refrigerated air for 167 d or more were severelydeteriorated, and thus not assessed for the aforementionedripening parameters. 2.3. Evaluation of physiological disorders The pears evaluated for quality were also evaluated for theincidence of senescent scald and internal breakdown, which wasexpressed as the percentage fruit displaying injury, regardless ofseverity, and averaged for the four treatment replicates. 2.4. Extraction and analysis of oxidative stress metabolites Unless otherwise mentioned, chemical reagents were pur-chased from Sigma-Aldrich (Oakville, ON, Canada). For eachcultivar, the GABA concentration per treatment replicate wasdetermined following extraction of pooled pulverized fruitpowders, representing equal amounts of four pear sub-samples,as described previously (Allan and Shelp, 2006; Lum et al.,2016a).Briefly,1 g of the pooled fine frozen powder was ground in 4 mL ofsulfosalicylic acid (30 mg mL-1) using a chilled mortar and pestleand fine silica sand. An aliquot (1.0 mL) of the homogenate was Fig. 1. Effect of 1-MCP and CA on the development of senescent scald in stored ‘Cold Snap’and'Swiss Bartlett'pear fruit. Fruit were treated with or without 1-MCP on the dayof harvest and stored at O°C under refrigerated air or CA of 18 or 2.5 kPa O2, and 2 kPa CO2. No 1-MCP (closed black squares); 1-MCP(open greysquares). Data represent themean incidence of four storage replicates of 10 fruit each. Shared letters at each time point or within a treatment indicate no significant difference at the P≤ 0.05 level. Capitalletters represent significant differences over storage time (within each treatment), whereas small letters represent significant differences among storage conditions (acomparison across all six treatments) at each time. Within each plot, black and grey letters correspond to the proximal non-1-MCP or 1-MCP datum, respectively. centrifuged and supernatant was neutralized with 4N NaOH. Theextract was passed through a 0.45 um syringe filter, followed byon-line o-phthalaldehyde derivatization and reverse-phase highperformance liquid chromatography. Whole fruit ascorbate andglutathione concentrations were determined for each treatmentreplicate following hydrochloric acid extraction of pooled pulver-ized fruit powder (100 mg in total) representing 25 mg each of fourfruit sub-samples essentially as described by Lum et al. (2016a).The neutralized extracts were used to estimate fruit AA, DHA, GSHand GSSG with enzyme-coupled microplate assays on a Spec-traMax Plus 384 spectrophotometer (Molecular Devices, Sunny-vale, CA, U.S.A.) according to previously published protocols (Lumet al., 2016a; Queval and Noctor, 2007). All neutralized extractswere assayed in duplicate. level. Replicate room effects were analyzed using ANOVAs (procmixed method) to partition variance into the fixed effects (1-MCP,CA regime and storage time) and their interactions, and therandom effect of chambers. In cases where interactions weresignificant (P≤0.05), treatment means were compared withineach sampling time (slice option) over the storage period using aFisher's protected least significant difference (LSD) test. Forexperiments where interactions were significant (P≤0.05), datafor each storage treatment replicate were pooled across repeatedmeasures to determine differences among treatments and overstorage period using Fisher’s protected LSD. 3. Results 2.5. Statistical analyses 3.1. Impact of 1-MCP, CA and low 02 on physiological disorders duringpostharvest storage 2.5.1. Analysis of variance (ANOVA) between treatment and withintreatment effects For fruit quality and metabolite data, statistical analyses wereconducted using SAS 9.3 (SAS Institute Inc., Cary,NC) at thea=0.05 The incidence of physiological disorders was compared for fruitof both cultivars, which were treated with or without 1-MCP andthen stored at 0°C under refrigerated air or CA (18 kPa O2, 2kPaCO2;2.5 kPa 02,2 kPa CO2). Senescent scald was absent or marginal Fig.2. Effect of 1-MCP and CA on the development of internal breakdown in stored 'Cold Snap'and 'Swiss Bartlett' pear fruit. Fruit were treated with or without 1-MCP on theday of harvest and stored at 0°Cunder refrigerated air or CA of 18 or 2.5 kPa O2,and 2 kPa C02. No 1-MCP(closed black squares); 1-MCP (open grey squares). Data represent themean incidence of four storage replicates of 10 fruit each. Shared letters at each time point or within a treatment indicate no significant difference at the P≤0.05 level. Capitalletters represent significant differences over storage time (within each treatment), whereas small letters represent significant differences among storage conditions (acomparison across all six treatments) at each time. Within each plot, black and grey letters correspond to the proximal non-1-MCP or 1-MCP datum, respectively. 'Cold Snap' Fig. 3. Effect of 1-MCP and CA on fruit quality parameters in 'Cold Snap’ and 'Swiss Bartlett’ pear fruit. Fruit were treated with or without 1-MCP on the day of harvest andstored at O°C under refrigerated air or CA of 18 or2.5 kPa O2, and 2 kPa CO2. No 1-MCP (closed black squares); 1-MCP(open grey squares). Ethylene production rates, firmnesslevels and surface peel colour were assessed at harvest and upon the removal of fruit from storage and transfer to 22 °C for up to 6 h. Peel colour was measured subjectively onthe following scale: 1=green, 2=more green than yellow, 3=more yellow than green, 4=yellow. For each quality parameter, data represent the mean of four storagereplicates of 10 fruit each. Shared letters at each time point or within a treatment indicate no significant difference at the P≤0.05 level. Capital letters represent significant (≤21%) for the first 111 to 119 d of most treatments (Fig. 1).Thereafter, approximately 60-100% incidence was observed innon-1-MCP fruit of both cultivars chilled under both refrigeratedair and 18 kPa O2; 'Swiss Bartlett’ displayed the lowest incidence ofthis injury at 18 kPa 02. With 2.5 kPa 02, the incidence wasmarginal, regardless of cultivar. Overall, senescent scald wasminimized by 1-MCP under refrigerated air and to a greater extentby CA, with the lowest levels being observed at 2.5 kPa 02. It isworth mentioning that symptoms of superficial scald were notobserved in pears of either cultivar stored under any treatment. A high incidence of internal breakdown was most evident innon-1-MCP‘Cold Snap'pears during prolonged storage, regardlessof whether CA was used; disorder levels were reduced by 39 to 89%with 1-MCP, and the most effective control was with 2.5 kPa O2(Fig. 2). Similarly, 1-MCP and CA dramatically limited thedevelopment of symptoms for internal breakdown in ‘SwissBartlett' pears in comparison to non-1-MCP pears held underrefrigerated air; disorder levels were reduced by 52 to 72% with CA,and nearly eliminated by 1-MCP, regardless of the CA regime used. 3.2. Impact of 1-MCP and CA on ripening capacity Although ‘Cold Snap’ fruit were harvested at a slightly lowerfirmness than ‘Swiss Bartlett'(representative of their respectivecommercial harvest maturities), firmness changed very little(≤15%loss) with storage(Fig. 3). Furthermore, the rate of softeningwas similar across all 1-MCP treatments,regardless of CA regime.By contrast, the rate of softening was greatest in non-1-MCP fruit ofthe ‘Swiss Bartlett'cultivar relative to the minimal losses (≤10%)observed in all 1-MCP fruit. The peel surface colour of ‘Cold Snap'fruit was mostly yellow after 119 d of refrigerated air storage,regardless of 1-MCP treatment. This phenomenon was delayedwhen 1-MCP fruit were held under CA, with the least yellowingapparent after 180 d at2.5 kPa O2 (Fig.3). In contrast, there was adramatic effect of 1-MCP and CA on peel yellowing in ‘SwissBartlett' fruit; specifically, this colour change was delayed by 8weeks in 1-MCP fruit stored under refrigerated air, and by 16 weeksunder CA. Peel colouration of 1-MCP fruit of ‘Swiss Bartlett'storedin 2.5 kPa O2 changed very little with storage duration. 3.3. Impact of 1-MCP, CA and low 02 on GABA concentrations Changes in whole-fruit GABA concentration as a function ofstorage period varied with cultivar, 1-MCP and storage atmosphere(Fig.4). The GABA concentration in 'Swiss Bartlett' fruit at harvest was 2.8 times higher than that of‘Cold Snap’fruit, and in both casesthere was no change after the pre-storage 1-MCP treatment.Moreover, GABA concentrations in Cold Snap'fruit were increasedby 2.5- to 4-fold within 1 week of the transfer to 0°C, whereasconcentrations in ‘Swiss Bartlett’ fruit decreased by 22-61%.Thereafter, GABA concentrations remained stable for the next 16weeks in all ‘Cold Snap'fruit stored under refrigerated air and the18 kPa 02 CA regime, whereas fluctuations were evident with2.5 kPa 02. By 180 d, the GABA concentrations increased by 1.4- to2.8-fold in all ‘Cold Snap'fruit stored under refrigerated air or innon-1-MCP fruit stored at 2.5 kPa O2; very little change wasapparent for all remaining treatments. In contrast, 1-MCPincreased the GABA concentrations in ‘Swiss Bartlett’fruit by36-190% after 34 d, compared to non-1-MCP fruit; concentrationsin 1-MCP fruit remained unchanged for the following 11 weeks, butdecreased thereafter. After 167 d, non-1-MCP fruit of ‘SwissBartlett' held under refrigerated air displayeda 15-fold increase inGABA concentration. No such increases were apparent for anyother treatment. 3.4. Impact of 1-MCP, CA and low 02 on antioxidant metabolism Ascorbate and glutathione were detected as reduced andoxidized forms (Figs. 5 and 6; Supplemental Figs. S1 and S2). Forboth cultivars, the composition of total ascorbate at harvest waspredominantly AA and gradually declined with storage duration. InCold Snap'fruit, total ascorbate levels declined by 64-76% after 28d of storage, with the smallest change observed in 1-MCP fruit at2.5 kPa O2 (Fig.5). A further 60% loss was evident by the end of thestorage period; final ascorbate levels were similar across alltreatments.These losses coincided with comparable changes in AAconcentrations as a function of storage period (SupplementalFig. S1). In contrast, the concentration of DHA and the AA/DHAratio for all treatments fluctuated within the first 56 d(Supplemental Fig. S1; Fig. 5). Only 1-MCP fruit held underrefrigerated air displayed a transient second spike in this ratio at119 d. By comparison, 1-MCP ‘Cold Snap’ fruit displayed minorfluctuations in AA/DHA at 2.5kPa 02 relative to all othertreatments. ‘Swiss Bartlett' fruit displayed a rapid decline in total ascorbateand the predominant form, AA, over the initial 34 d, regardless oftreatment (Fig. 5; Supplemental Fig. S1). Interestingly, theseconcentrations were maintained beyond 34 d in all 1-MCP fruitrelative to non-1-MCP fruit. Fluctuations in DHA concentration andAA/DHA redox status were apparent in all treatments throughoutthe storage period (Fig. 5; Supplemental Fig. S1). With prolongedstorage, DHA levels tended to be higher with 1-MCP than without1-MCP. Interestingly, the AA/DHA ratio recovered to pre-storagelevels at 167 d in 1-MCP fruit maintained under refrigerated air andall fruit held at 2.5kPa O2. For both cultivars, fluctuations in fruit glutathione levels andGSH/GSSG ratios were apparent with storage period. With alltreatments, the total glutathione concentrations declined inColdSnap’fruit over the initial 7 d of storage, although minor spikeswere apparent thereafter (Fig. 6). All fruit stored under refrigeratedair displayed the largest drop in total glutathione levels after 180 dof storage. By comparison, slightly lower decreases in totalglutathione concentration were evident for all fruit subjected toCA. Simultaneous and similar changes in GSH concentrations werefound in all treatments as a function of storage period (Supple-mental Fig. S2). Conversely, a rapid and 100% increase in GSSGlevels occurred by 7 d of storage, but fluctuated thereafterregardless of 1-MCP and storage regime (Supplemental Fig. S2). Fig. 4. Effect of 1-MCP and CA on GABA concentration in stored ‘Cold Snap'and'Swiss Bartlett' pear fruit. Fruit were treated with or without 1-MCP on the day of harvest and. stored at 0°C under refrigerated air or CA of 18 or 2.5 kPa O2, and 2 kPa CO2. No 1-MCP (closed black squares); 1-MCP (open grey squares). Data represent the mean GABAconcentration (in pmolkg-) of four storage replicates of four fruit each. The initial datum at 0 minus 24 h and O represents fruit at harvest and following 1-MCP treatment,respectively. Shared letters at each time point or within a treatment indicate no significant difference at the P≤0.05 level. Capital letters represent significant differences overstorage time (within each treatment), whereas small letters represent significant differences among storage conditions (a comparison across all six treatments) at each timeWithin each plot, black and grey letters correspond to the proximal non-1-MCP or 1-MCP datum, respectively. At the end of the experiment, 1-MCP fruit maintained under CAcontained the highestconcentrations of GSSG. For all treatments, arapid and transient decline (i.e., 80%) in GSH/GSSG occurred upontransfer to low temperature (Fig. 6). A temporary recovery of thisratio was evident after 28 d in fruit stored under refrigerated airand in non-1-MCP fruit at 2.5 kPa O2; smaller ratios were apparentfor the remaining treatments. After 180 d of storage, the final ratioof GSH/GSSG was in the range of 0.21 to 1.86, with the latterapparent for 1-MCP fruit stored under 18 kPa O2; on the whole thisratio was markedly lower than pre-storage levels. 'Swiss Bartlett’ fruit exposed to CA displayed a 28% decline intotal glutathione levels during the first6 d of storage,regardless of1-MCP treatment (Fig. 6). This decline was delayed by a week in allfruit held under refrigerated air. Thereafter, transient increases intotal glutathione were evident in 1-MCP fruit stored at 18 kPa 02and all fruit held under refrigerated air. Marginal changes wereevident relative to pre-storage levels for 1-MCP fruit stored under 2.5 kPa O2. Interestingly, at 167 d total glutathione levels in 1-MCPfruit approximated pre-storage concentrations, regardless ofatmospheric conditions; in contrast, total glutathione concen-trations in non-1-MCP fruit were 20 to 33% of those detected atharvest. Similar changes were apparent for GSH concentration as afunction of storage period (Supplemental Fig. S2). Conversely,GSSG levels were enriched by 1- to 2-fold within 13 d of storageand declined thereafter. A 76% decline in the GSH/GSSG ratio wasapparent after only 13 d of storage, regardless of CA regimes, andfluctuations in this ratio were apparent thereafter (Fig. 6). Ingeneral, concentrations of GSSG and GSH/GSSG ratios tended to behigher in 1-MCP fruit than non-1-MCP fruit. After 167 d of storage,the final GSH/GSSG ratio in 1-MCP fruit at 18 kPa O2 matched thepre-storage ratio. At this time, somewhat lower ratios were foundin 1-MCP fruit under the remaining storage regimes. The GSH/GSSG ratio was less than one for all non-1-MCP fruit of ‘SwissBartlett’ by the end of the storage period. 'Cold Snap' 600 [0℃, Refrigerated air [0C, Refrigerated air 一一No 1-MCP —NNo 1-MCP 8 Aa 1-MCP 一口-1-MCP 450 6 -a 300 4 ga .Abc Bab 150一 Bab Babc: Ba BCa 2 a BCab Db Da CDa BCab DEab Da 0 LD 600 .0℃, 18 kPa O,, 2 kPa Co, 0℃, 18 kPa O,, 2 kPa CO, 8Fo ou 450 300 Aab 6工 4 ABa 150 Bb BCa BCab 2 F Bc BCa Bb ABa Ba Db CDa 0 lBd 0 600 0℃, 2.5 kPa O,2 kPa Co, 0℃, 2.5 kPa 0,2 kPa CO, 8 450 6 Aa 300 Aab 4 ABbc tAbc Baa 150 Babc Ca CDab 2 ABbBCa Da 3b Ba Cb cDa Db CDal CDab ABd 0 0)L CDb. 5 10 15 20 25 0 5 10 15 20 25 Storage period (weeks) Storage period (weeks) .'Swiss Bartlett'. Fig.5. Effect of 1-MCP and CA on concentrations of total ascorbate and ratio of AA/DHA in stored ‘Cold Snap'and'Swiss Bartlett' pear fruit. Fruit were treated with or without1-MCP on the day of harvest and stored atO°Cunder refrigerated air or CA with 18 or 2.5 kPa O2, and 2 kPa C02. No 1-MCP (closed black squares); 1-MCP (open grey squares).Data represent the mean ascorbate concentration (in umolkg)or ratio of four storage replicates of four fruit each. The initial datum at 0 minus 24 h and O represents fruit atharvest and following 1-MCP treatment, respectively. Shared letters at each time point or within a treatment indicate no significant difference at the P≤ 0.05 level. Capitalletters represent significant differences over storage time (within each treatment), whereas small letters represent significant differences among storage conditions (acomparison across all six treatments) at each time. Within each plot, black and grey letters correspond to the proximal non-1-MCP or 1-MCP datum, respectively. 0℃, Refrigerated air 0℃, Refrigerated air —— No 1-MCP —— No 1-MCP Storage period (weeks) Storage period (weeks) Fig. 6. Effect of 1-MCP and CA on concentrations of total glutathione and ratio of GSH/GSSG in stored‘Cold Snap’and 'Swiss Bartlett’ pear fruit. Fruit were treated with orwithout 1-MCP on the day of harvest and stored at 0°C under refrigerated air or CA with 18 or2.5 kPa O2, and 2 kPa CO2. No 1-MCP (closed black squares); 1-MCP (open greysquares). Data represent the mean glutathione concentration (in umolkg) or ratio of four storage replicates of four fruit each. The initial datum at 0 minus 24 h and Orepresents fruit at harvest and following 1-MCP treatment, respectively. Shared letters at each time point or within a treatment indicate no significant difference at theP≤ 0.05 level. Capital letters represent significant differences over storage time (within each treatment), whereas small letters represent significant differences among 4.1.1-MCP and CA inhibited the development of senescent scald andinternal breakdown In this study, internal breakdown and senescent scald wereabsent over the first 111 to 119 d of storage in fruit of'Swiss Bartlett'and Cold Snap’,respectively (Figs. 1 and 2). Physiological injuriesdeveloped thereafter in both cultivars, and these were mostevident under refrigerated air storage when 1-MCP was notapplied prior to storage. These disorders are known to occur inpears of other cultivars during prolonged storage (i.e., beyond threemonths) (Franck et al., 2007; Kupferman, 2003). Over the lastdecade, various studies of European pears have indicated that CAeffectively controls these disorders, thereby extending theirstorage life (Drake and Chen, 2000; Drake and Elfving, 2004;Lum et al., 2016b). Alternatively, 1-MCP minimizes these disordersin ‘Bartlett’fruit (DeEll and Ehsani-Moghaddam, 2011; Ekmanet al., 2004; Villalobos-Acuna et al., 2011a; Wang and Sugar, 2015).In our study, the incidences of both senescent scald and internalbreakdown were correlated to 02 partial pressure in the storageatmosphere (i.e., disorders in refrigerated air >18 kPa O2>2.5 kPaO2; Figs.1 and 2); moreover, greater inhibition was evident with acombination of CA and 1-MCP. The most effective treatment forcontrol of senescent scald and internal breakdown in ‘Cold Snap’and ‘Swiss Bartlett’ fruit was 2.5kPa 02/2kPa CO2. UnlikeConference'pears (Franck et al., 2007; Lammertyn et al., 2003),low 02 disorders were not apparent in the ‘Bartlett’relatedcultivars investigated in this study. 4.2. Control ofethylene-mediated ripening was more dramatic in fruitof 'Swiss Bartlett' than Cold Snap' 1-MCP fruit of both cultivars displayed lower rates of ethyleneproduction following their removal from storage than control fruit(Fig. 3).Moreover, CA acted synergistically with 1-MCP to furtherreduce ethylene production in 'Cold Snap'and'Swiss Bartlett' fruitupon their removal from storage. This is not without precedent as‘Delicious', ‘Empire'and ‘Law Rome'apples exposed to 1-MCP andsubjected to CA storage (0C, 2.5kPa O2, 2.5 kPa CO2) for up to 32weeks produce much less ethylene than non-1-MCP fruit (DeEllet al., 2007; Watkins et al., 2000). The control of storage-relatedphysiological disorders by 1-MCP in ‘Bartlett’ fruit tends tocoincide with delayed and/or lower ethylene emission and reducedfruit softening (DeEll and Ehsani-Moghaddam, 2011; Ekman et al.,2004; Villalobos-Acuna et al., 2011a, 2011b). In our study, themaximal rates of ethylene production in non-1-MCP fruit storedunder refrigerated air tended to occur several weeks in advance ofdisorders in both cultivars. In contrast, similar rates of ethyleneproduction occurred in non-1-MCP fruit of ‘Swiss Bartlett'stored at2.5 kPa O2, but these fruit were devoid of disorders. Thus, there isno conclusive evidence that the capacity to produce ethylenepredisposes these fruit to storage-related disorders. The transition of peel colour from green to yellow in Europeanpears occurs with ripening, including in response to short-termchilling (i.e., four months; DeEll and Ehsani-Moghaddam, 2011;Villalobos-Acufia and Mitcham, 2008). In our study, peel yellowingwas evident during low temperature storage, although there werecultivar-specific differences with respect to the effect of CA and 1-MCP (Fig.3). Unlike ‘Swiss Bartlett’ fruit, there was very littleinhibition of peel yellowing in ‘Cold Snap'fruit by 1-MCP and CA,which could be associated with their relatively lower rates ofethylene production.The control of peel yellowing by 1-MCP in 'Swiss Bartlett'fruit is consistent with its beneficial effect on lowtemperature-stored‘Bartlett' fruit (DeEll and Ehsani-Moghaddam,2011; Ekman et al., 2004; Villalobos-Acuna et al., 2011a). Inaddition, 1-MCP was most effective in delaying this colourtransition in 'Swiss Bartlett' fruit under the 2.5 kPa 02/2kPa CO2regime. This is consistent with previous research demonstratingthat low 02 in combination with elevated CO effectively limitspeel yellowing in ‘Anjou’ pears (Drake, 1994). In general, fruit softening in pear fruit is dependent uponethylene biosynthesis (Hiwasa et al., 2003). Indeed, in our studysoftening was evident in non-1-MCP-fruit of‘Swiss Bartlett', whichwere shown to be capable of producing ethylene; little softeningwas evident in all 1-MCP fruit, which coincided with a low capacityto produce ethylene (Fig. 3). However, losses in firmness of ‘ColdSnap' fruit were only 9 to 15% throughout the storage period, andmore importantly these were not influenced by 1-MCP and CA.These findings are consistent with stable levels of firmness whichare found in ripening Chinese and Japanese pears, and associatedwith limited activity of the cell wall modifying enzyme endo-polygalacturonase (Hiwasa et al., 2004). This suggests that fruitsoftening and the production of endogenous ethylene in ColdSnap’ fruit are independent processes. 4.3. GABA accumulation was moderately associated with internalbreakdown and senescent scald In the present study, we hypothesized that the development ofsenescence-related disorders in ‘Cold Snap’and ‘Swiss Bartlett'fruit is associated with the accumulation of oxidative stressmetabolites during storage. GABA accumulated in stored fruit,although these increases were not exclusively associated with theoccurrence of senescent scald and internal breakdown, regardlessof pear cultivar (Figs. 1, 2 and 4). Previous research showed thatsoggy breakdown and CA-related injury in ‘Honeycrisp'apple fruitcoincide with enhanced GABA levels (Leisso et al., 2015; Lum et al.,2016a). Moreover, GABA is relatively more abundant in browncortical tissue than unaffected tissue of Conference'pears duringprolonged storage under 10 kPa CO2 and 1 kPa O2 (Franck et al.,2007; Pedreschi et al., 2009). On the whole, the absence of adistinct relationship between GABA and senescence-relateddisorders in Cold Snap’ and ‘Swiss Bartlett’pear fruit could beexplained by the elevation of GABA levels, albeit to varyingdegrees, in both disordered and symptomless fruit (Fig. 4). Inmelon, there is an increasing GABA gradient towards the center ofthe fruit; the lowest concentrations are found in the fruit epicarpand green mesocarp (Biais et al., 2010). It is not known whether aGABA gradient exists in stored pear fruit; in our study metaboliteanalysis was performed on whole fruit powder as GABA metabo-lism may be altered by mechanical handling (Shelp et al., 2012a). At the biochemical level,GABA accumulation in disordered fruitof Cold Snap'and ‘Swiss Bartlett' is most likely due to cytosolicacidification-mediated stimulation of glutamate decarboxylaseactivity, which is postulated to occur with cellular breakdown(Lum et al., 2016b). It is also possible that damage from ROSproduction results in mitochondrial membrane disintegration,allowing the release of Cafrom mitochondria and subsequentlyenhancing GABA production via the Ca2+/calmodulin activation ofglutamate decarboxylase (Shelp et al., 2012a, 2012b). In fact,evidence exists for both calmodulin-dependent and -independentglutamate decarboxylases in‘Empire' apple fruit (Trobacher et al.,2013). To date, it is not known if similar biochemical regulatoryprocesses operate within pear fruit, including those investigated inthis study. In addition, GABA levels in stored Cold Snap'and 'Swiss Bartlett’fruit could be due in part to catabolic processes. GABAlevels declined in 1-MCP fruit of 'Swiss Bartlett' with prolonged CAstorage, implyinga greater rate of GABA degradation than the rateof GABA production (Fig.4). It is widely accepted that catabolism ofGABA is diverted, at least to some extent, from the production ofsuccinate to y-hydroxybutyrate when the NADPH/NADP*ratio isaltered (Lum et al., 2016b;Shelp et al., 2012a). 4.4. Internal breakdown and senescent scald were associated with adecline in glutathione levels In this study, total ascorbate and AA concentrations declinedwith storage in fruit of both cultivars (Fig.5). The levels present atharvest and in stored fruit approximated those detected in otherEuropean pear cultivars when expressed on a umol kg- freshmass basis (Cascia et al., 2013). With ‘Swiss Bartlett', totalascorbate and AA levels were better maintained in 1-MCP fruitthan their non-1-MCP counterparts. A similar benefit of 1-MCPoccurs in ‘Conference’ and ‘Passe Crassane’ pears during lowtemperature storage (Chiriboga et al., 2013; Cocetta et al., 2016).The lower level of ascorbate in non-1-MCP fruit of 'Swiss Bartlett'could be somewhat related to their advanced ripened state andgreater capacity to produce ethylene. Previous research has shownthat exogenous ethylene decreases AA levels in spinach leaves, andascorbate losses are less rapid in ethylene-insensitive mutants ofArabidopsis than wild-type plants (Gergoff et al., 2010). In stressed plant tissues, a portion of the AA pool can beinterconverted to DHA (Foyer and Noctor, 2011). Both ‘Cold Snap’and 'Swiss Bartlett' fruit displayed transient increases in the levelsof DHA during storage (Supplemental Fig. S1). These shifts in DHAlevels,together with the limited maintenance of AA pools in stored'Cold Snap'and non-1-MCP fruit of'Swiss Bartlett' (Fig. 5), could beindicative of a dysfunctional AA-GSH recycling pathway. Forexample, a spike in DHA levels in ‘Williams Bon Chretien’ pearsduring this initial 3 d of CA storage appears to be associated withdecreased gene expression for the ascorbate recycling enzymesDHA reductase and monodehydroascorbate reductase (Casciaet al., 2013). Moreover, the transient accumulation in DHAconcentration has been linked to flesh browning in WilliamsBon Chretien’pears, but not in ‘Beurre Bosc’and ‘Doyenne duComice’pears. Our study demonstrated that changes in theconcentrations of DHA and AA occurred in advance of theappearance of senescence-related disorders in Cold Snap’ and‘Swiss Bartlett' pear fruit (Figs. 1, 2 and Supplemental Fig. S1).Other research has shown that CA-related injury in ‘Honeycrisp'apple fruit is well associated with declining glutathione levels andglutathione redox status, but not ascorbate depletion (Lum et al.,2016a). For the most part, 1-MCP fruit of both cultivars displayed highertotal glutathione levels than non-1-MCP fruit during storage,although this difference was more evident with ‘Swiss Bartlett'(Fig. 6). To our knowledge, this is the first study demonstrating aneffect of 1-MCP on glutathione concentrations in stored pear fruit.Moreover, high total glutathione and GSH concentrations in 1-MCPfruit of ‘Swiss Bartlett'coincided with relatively more ascorbatepreservation and negligible disorder symptoms during prolongedstorage than non-treated fruit. Notwithstanding, in both cultivarsinternal breakdown and senescent scald development werepreceded by a decrease in the levels of total glutathione andGSH(Figs. 1, 2, and 6), implying that a decline in these levels renderthese fruit susceptible to these disorders. The findings reportedhere are inconsistent with the accumulation of total glutathione inbrown-core tissue of 'Blanquilla' pears (Pinto et al., 2001). Thisdiscrepancy could be due to our use of whole-fruit glutathioneconcentrations as a function of storage period, rather thandissected tissues of fruit sampled at the end of the storage experiment as done previously. By far, the most rapid loss of totalglutathione was in non-1-MCP pears maintained under refrigerat-ed air, a treatment combination that culminated in the greatestcapacity to produce ethylene and the highest incidence ofsenescence-related disorders. Together, these findings suggestthat there is a possible link between glutathione and ethyleneproduction in stored ‘Cold Snap'and ‘Swiss Bartlett'pear fruit. Invegetative tissues of Arabidopsis, GSH regulates ethylene biosyn-thesis and the role of this phytohormone in the resistance to saltstress (Datta et al., 2015). Furthermore, ethylene action is requiredfor glutathione-mediated control of ozone-related oxidative stressin Arabidopsis (Yoshida et al., 2009). To date, it is unknownwhether there is any cross-talk between ethylene and glutathionemetabolism in stored pear fruit. 5. Concluding remarks Prior to this study, there was little information on the impact ofCA and 1-MCP on the quality and senescence of'Swiss Bartlett'and‘Cold Snap’ pear fruit during low temperature storage. Senescentscald and internal breakdown occurred simultaneously during lowtemperature storage of both cultivars under consideration, andsymptoms were most prominent with a combination of non-1-MCP and refrigerated air. 1-MCP in combination with elevated CO,storage effectively controlled these disorders in Cold Snap' and‘Swiss Bartlett' pears. An analysis of oxidative stress metaboliteprofiles revealed that GABA levels were enhanced with storage,although there was no consistent effect of 1-MCP and CA. Cultivar-specific differences were observed with respect to changes inascorbate levels; 1-MCP tended to preserve total and reducedascorbate concentrations in 'Swiss Bartlett'fruit across all storageatmospheres. In fruit of both cultivars, total and reducedglutathione levels declined and the GSH/GSSG ratio fluctuatedwith storage. In general, these levels were better maintained in 1-MCP fruit. Overall, it appears that 1-MCP in combination with low02 limits senescence disorders during prolonged low temperaturestorage. Acknowledgements Research for this project was supported by funding awardedtoGGB and BJS from the Natural Sciences and Engineering ResearchCouncil (NSERC) of Canada as a Collaborative Research andDevelopment Grant, AgroFresh Inc., the Niagara Peninsula Fruitand Vegetable Growers’Association, and the Ontario Ministry ofAgriculture, Food and Rural Affairs. We are grateful to BehrouzEhsani-Moghaddam for technical assistance with analysis of fruitripening indices. Appendix A. Supplementary data Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.postharvbio.2017.03.008. References Allan, W.L., Shelp, B.J., 2006. Fluctuations of y-aminobutyrate, y-hydroxybutyrate,and related amino acids in Arabidopsis leaves as a function of the light-darkcycle,leaf age, andN stress. Can. J. Bot. 84, 1339-1346. Argenta, L.C., Mattheis, J.P., Fan, X., Amarante, C.V.T., 2016. Managing'Bartlett'pearfruit ripening with 1-methylcyclopropene reapplication during cold storage.Postharvest Biol. Technol. 113,125-130. Biais, B., Beauvoit, B., Allwood, J.W., Deborde, C., Maucourt, M., Goodacre, R., Rolin,D., Moing, A., 2010.Metabolic acclimation to hypoxia revealed by metabolite?-?4gradients in melon fruit. J. Plant Physiol. 167, 242-245. Bonn, W.G., van der Zweet, T., 2000. Distribution and economic importance of fireblight. In: Vanneste, J.L. (Ed.), Fire Blight: the Disease and its Causative Agent,Erwinia amylovora. CAB International Publishing, New York, pp. 37-53. Cascia, G., Bulley, S.M., Punter, M., Bowen, J., Rassam, M., Schotsmans, W.C.,Larrigaudiere, C., Johnston, J.W., 2013. Investigation of ascorbate metabolismduring inducement of storage disorders in pear. Physiol. Plant. 147, 121-134. Chiriboga,M.A., Bordonaba,J.G.,Schotsmans,W.C., Larrigaudiere Recasens, I., 2013. Antioxidant potential of 'Conference' pears during cold storage and shelf life inresponse to 1-methylcyclopropene. Lebenson Wiss Technol. 51, 170-176. Cocetta,G., Mignani, I., Spinardi, A., 2016. Ascorbic acid content in ‘Passe-Crassane’winter pear as affected by 1-methylcyclopropene during cold storage and shelflife. HortScience 51, 543-548. Datta, R., Kumar, D., Sultana,A., Hazra,S.,Bhattacharyya,D., Chattopadhyay,S., 2015.Glutathione regulates 1-aminocyclopropane-1-carboxylate synthasetranscription via WRKY33 and 1-aminocyclopropane-1-carboxylate oxidase bymodulating messenger RNA stability to induce ethylene synthesis during stress.Plant Physiol. 169, 2963-2981. DeEll, J.R., Ehsani-Moghaddam, B., 2011. Timing of postharvest 1-methylcyclopropene treatment affects Bartlett pear quality after storage. Can. J.Plant Sci. 91.853-858. DeEll,J.R.,Ayers,J.T., Murr, D.P., 2007.1-Methylcyclopropene influences ‘Empire' and'Delicious' apple quality during long-term commercial storage. HortTechnology17.46-51. Deuchande, T., Larrigaudiere, C., Gine'-Bordonaba,J., Carvalho, S.M.P., Vasconcelos,M.W.,2016.Biochemical basis of COz-related internal browning disorders inpears (Pyrus communis L. cv. Rocha) during long-term storage.J. Agric. FoodChem.64,4336-4345. Drake, S.R., Chen, P.M., 2000. Storage quality of ethylene treated 'd'Anjou'and 'Bosc'winter pears. J. Food Process. Preserv. 24, 379-388. Drake, S.R., Elfving, D.C., 2004. Quality of packed and bin stored ‘Anjou' pears asinfluenced by storage atmosphere and temperature. J. Food Qual. 27, 141-152. Drake, S.R., 1994. Elevated carbon dioxide storage of 'Anjou' pears using purge-controlled atmosphere. HortScience 29, 299-301. Ekman, J.H., Clayton, M., Biasi, W.V., Mitcham, EJ., 2004. Interactions between 1-MCP concentration, treatment interval and storage time for ‘Bartlett'pears.Postharvest Biol. Technol.31, 127-136. FAOSTAT, 2016. database. Food and Agriculture Organization of the United Nations,Statistics Division, Rome. http://faostat3.fao.org/home/E. Accessed 29 Nov2016. Foyer, C.H., Noctor, G., 2011. Ascorbate and glutathione: the heart of the redox hub.Plant Physiol.155,2-18. Franck, C., Lammertyn, J.,Ho, Q.T., Verboven, P., Verlinden, B., Nicolai, B.M., 2007.Browning disorders in pear fruit. Postharvest Biol. Technol. 43,1-13. Gago, C.M.L., Miguel, M.G., Cavaco, A.M., Almeida, D.P.F., Antunes, M.D.C., 2015. Combined effect of temperature and controlled atmosphere on storage andshelf-life of‘Rocha'pear treated with 1-methylcyclopropene. Food Sci. Technol.Int. 21. 94-103. Gergoff, G., Chaves, A., Bartoli, C.G., 2010. Ethylene regulates ascorbic acid contentduring dark-induced leaf senescence. Plant Sci. 178, 207-212.Harris, R., 2016. Pear-fection. Can. Groc. Mag. 130, 32-35. Hiwasa, K., Kinugasa, Y., Amano, S., Hashimoto, A., Nakano, R., Inaba, A., Kubo, Y.,2003. Ethylene is required for both the initiation and progression of softening inpear (Pyrus communis L.) fruit. J. Exp. Bot. 54, 771-779. Hiwasa, K.,Nakano,R., Hashimoto,A., Matsuzaki, M., Murayama, H., Inaba, A., Kubo,Y.,2004.European, Chinese and Japanese pear fruits exhibit differentialsoftening characteristics during ripening. J. Exp. Bot.55, 2281-2290. Hunter, D.M., Slingerland, K.C., 2008. Evaluation of four fire blight-tolerant pearcultivars and selections for commercial pear production in Canada. Acta Hortic.800,541-546. Hunter, D.M., Kappel,F.,Quamme,H.A.,Bonn, W.G., Slingerland,K.C.,2009."HarovinSundown'pear. HortScience 44, 1461-1463. Ingels, C.A., Burkhart, D.J., Elkins, R.B., 2007. Varieties. In: Mitcham, E.J., Elkins, R.B.(Eds.), Pear Production and Handling Manual, 3483. Univ. Calif. Agr. Nat.Resources Publ., pp. 25-31. Kader, A.A., 2007. Controlled atmospheres. In: Mitcham,E.J., Elkins, R.B. (Eds.), PearProduction and Handling Manual, 3483. Univ.Calif. Agr. Nat. Resources Publ.,pp.175-177. Kupferman,E., 2003. Controlled atmosphere storage of apples and pears. Acta Hort.600,729-735. ( L a mmer t yn,J . , D r es sel a e r s, T., Van H e c ke, P., Jancsok, P . , We v er s , M . , N i co l ai, B.M. , 2 0 03. An a ly s i s of t he t i me co urse o f core b rea kdown in C o n feren c e p e ar s b y m e a ns o n M R I an d X-ray C T . P o s th a r vest Bi o l. Techn ol. 2 9 , 1 9 - 2 8 . ) Leisso, R.S., Buchanan,D.A., Lee,J.,Mattheis, J.P., Sater, C., Hanrahan, I., Watkins, C.B.,Gapper,N.,Johnston,J.W.,Schaffer, R.J., Hertog, M.L.A.T.M., Nicolai, B.M.,Rudell,D.R., 2015.Chilling-related cell damage of apple (Malus x domestica Borkh.) fruitcortical tissue impacts antioxidants, lipid and phenolic metabolism. Physiol.Plant.153,204-220. Lum, G.B., Brikis, C.J., Deyman, K.L., Subedi,S., DeEll, J.R., Shelp, B.J., Bozzo, G.G.,2016a. Pre-storage conditioning ameliorates the negative impact of 1-methylcyclopropene on physiological injury and modifies the response ofantioxidants and gamma-aminobutyrate in'Honeycrisp' apples exposed tocontrolled-atmosphere conditions. Postharvest Biol. Technol. 116, 115-128. Lum, G.B., Shelp, B.J., DeEll, J.R., Bozzo, G.G., 2016b. Oxidative metabolism isassociated with physiological disorders in fruits stored under multipleenvironmental stresses. Plant Sci. 245, 143-152. Lurie, S., Watkins, C.B., 2012. Superficial scald, its etiology and control. PostharvestBiol. Technol. 65, 44-60. Ma, S.S., Chen, P.M., 2003. Storage disorder and ripening behavior of 'Doyenne duComice’pears in relation to storage conditions. Postharvest Biol.Technol. 28,281-294. Pedreschi, R., Franck, C.,Lammertyn,J., Erban, A., Kopka,J., Hertog, M.,Verlinden, B.,Nicolai, B., 2009. Metabolic profiling of Conference' pears under low oxygenstress. Postharvest Biol. Technol. 51.123-130. Pinto, E.,Lentheric, I., Vendrell, M., Larrigaudiere, C., 2001. Role of fermentative andantioxidant metabolisms in the induction of core browning in controlled-atmosphere stored pears. J. Sci. Food Agric. 81, 364-370. Queval,G., Noctor, G., 2007.A plate reader method for the measurement of NAD,NADP, glutathione, and ascorbate in tissue extracts: application to redoxprofiling during Arabidopsis rosette development. Anal. Biochem. 363,58-69. Shelp, B.J., Bozzo, G.G., Trobacher, C.P., Chiu, G., Bajwa, V.S.,2012a. Strategies andtools for studying the metabolism and function of y-aminobutyrate in plants. I.Pathway structure. Botany 90, 651-668. Shelp, B.J., Bozzo, G.G., Zarei, A., Simpson, J.P., Trobacher, C.P., Allan, W.L., 2012b.Strategies and tools for studying the metabolism and function ofy-aminobutyrate in plants. II. Integrated analysis. Botany 90, 781-793. Trobacher, C.P., Zarei, A., Liu,J., Clark, S.M., Bozzo, G.G., Shelp, B.J., 2013. Calmodulin-dependent and calmodulin-independent glutamate decarboxylases in applefruit. BMC Plant Biol. 13, 144. Vanoli, M., Grassi, M., Eccher Zerbini, P., Rizzolo, A., 2010. Fluorescence, conjugatedtrienes, a-farnesene and storage disorders in ‘Abbé Fétel' pears cooled withdifferent speeds and treated with 1-MCP. Acta Hort. 858, 191-198. Villalobos-Acuna,M., Mitcham, E.J., 2008. Ripening of European pears: the chillingdilemma. Postharvest Biol. Technol. 49, 187-200.cz Villalobos-Acufia, M.G., Biasi, W.V., Flores, S., Jiang, C.Z., Reid, M.S., Willits, N.H., Mitcham, E.J., 2011a. Effect of maturity and cold storage on ethylenebiosynthesis and ripening in 'Bartlett'pears treated after harvest with 1-MCP.Postharvest Biol. Technol.59.1-9. Villalobos-Acufia, M., Biasi, W.V.,Mitcham, E.J., Holcroft, D., 2011b. Fruit temperature and ethylene modulate 1-MCP response in 'Bartlett'pears. Postharvest Biol. Technol. 60,17-23. Wang, Y., Sugar,D.,2015.1-MCP efficacy in extending storage life of‘Bartlett'pears isaffected by harvest maturity, production elevation, and holding temperatureduring treatment delay. Postharvest Biol. Technol. 103, 1-8. Watkins,C.B., Nock, J.F., Whitaker, B.D., 2000. Responses of early, mid and lateseason apple cultivars to postharvest application of 1-methylcyclopropene (1-MCP) under air and controlled atmosphere storage conditions.Postharvest Biol.Technol. 19, 17-32. Watkins, C.B., 2006. The use of 1-methylcyclopropene (1-MCP) on fruits andvegetables. Biotechnol. Adv.24, 389-409. Watkins, C.B., 2015.Advances in the use of 1-MCP. In:Willis, R.B.H.,Golding,J.(Eds.),Advances in Postharvest Fruit and Vegetable Technology. CRC Press, Boca Raton,FL, pp. 117-145. ( Y osh i d a , S., T amaoki, M., Io ki, M. , Ogawa,D. , S a t o,Y.,Aono , M . , Ku bo, A ., S aj i, S ., S aji , H . , S ato h,S . , N a k ajima , N . , 2 0 09 . E t h yle n e an d s alicyl ic a c id c ontr ol glu tat hi one b io sy n th es is in ozo n e-expo s ed A r ab i dops is t h a l ia n a. Phy s i ol . P l an t 136,2 84 -298 . Z ho u , M., Ndeuru mi o , K . H ., Zhao, L . , H u , Z., 2 0 1 6. Im p act o f pre c o o ling an d ) ( c o n tr o lle d - a tm osph er e s tor a ge o n y -a mi no but y r i c a c id (G ABA) a cc um ulat ion i n lo n ga n ( D im o c arpus lon g an Lo u r.) frui t. J . A g r i c . Food Chem. 6 4,6 4 4 3-6 4 5 0. ) http://dx.doi.org/j.postharvbio. differences over storage time (within each treatment), whereas small letters represent significant differences among storage conditions (a comparison across all sixtreatments) at each time. Within each plot, black and grey letters correspond to the proximal non--MCP or -MCP datum, respectively. 欧洲梨(Pyrus communis L.)在低温下贮藏以延长采后寿命。不幸的是,长时间暴露在贮藏环境中很可能发生老化烫伤和内部破裂。1-MCP能控制成熟障碍。本研究的主要目的是研究1-MCP和CA气调对冷藏‘Cold Snap’和‘Swiss Bartlett’梨果实品质的影响,包括生理障碍和氧化应激代谢产物。用或不加1-MCP处理鲜采梨,然后在0℃冷藏空气或CA气调(18kpa或2.5kpa氧气和2kpa CO2)下贮藏至少167d。1-MCP和CA延缓和/或降低了两个品种贮藏果实中乙烯的产生速率。1-MCP和CA延缓了‘Swiss Bartlett’梨果实软化和果皮变黄,但对‘Cold Snap’影响可忽略至轻微。在这两个品种中,未经1-MCP处理的梨在冷藏贮藏过程中发生衰老烫伤和内部破裂的发生率都很高。在大多数情况下,这些症状通过CA气调和1-MCP得以缓解,在2.5 kPa的氧气条件下,1-MCP处理的梨的发病率最低甚至可以忽略不计。g-氨基丁酸盐在贮藏梨中积累,尽管1-MCP和CA稍微降低了“Cold Snap”水果中的含量,而1-MCP在“瑞士巴特利特”水果中增加了含量。抗坏血酸(总含量和还原量)水平在“寒流”水果中迅速耗尽,无论处理方式如何;在所有贮藏环境中,1-MCP处理的“Swiss Bartlett”水果比对照水果的水平保持得更好。在两个品种中,谷胱甘肽(总的和还原的)浓度和氧化还原状态在贮藏期间都有波动,尽管1-MCP处理的果实中这些水平普遍较高。此外,贮藏梨果实中谷胱甘肽缺乏发生在衰老障碍发生之前
确定






还剩10页未读,是否继续阅读?

产品配置单


图拉扬科技有限公司为您提供《梨中CO2,硬度,色差,糖酸度检测方案(气调保鲜)》,该方案主要用于其他水果制品中理化分析检测,参考标准--,《梨中CO2,硬度,色差,糖酸度检测方案(气调保鲜)》用到的仪器有Labpod Mini气调保鲜试验系统、Safepod DCA 气调保鲜系统
推荐专场






相关方案
更多
该厂商其他方案
更多