








方案详情
文
在植物-细菌生物炭系统模型中芘的生物转化和渗透过程中的风险降低Risk reductions during pyrene biotransformation and mobilization in a model plant-bacteria-biochar system
采用格哈特公司 索克森Soxtherm 全自动快速固液萃取系统 基于固液萃取原理使用100 mL的丙酮/二氯甲烷(1:1v/v)测定5g选定渗滤液中12c-芘的残余浓度。
The residual concentration of 12C-pyrene was also measured in selected leachate samples by exhaustive solid-liquid extraction using a subsample of 5 g and a mixture of 100 mL of acetone/dichloromethane (1:1 v/v) with a Soxtherm extractor (C. Gerhardt GmbH & Co. KG, Königswinter,Germany).
方案详情

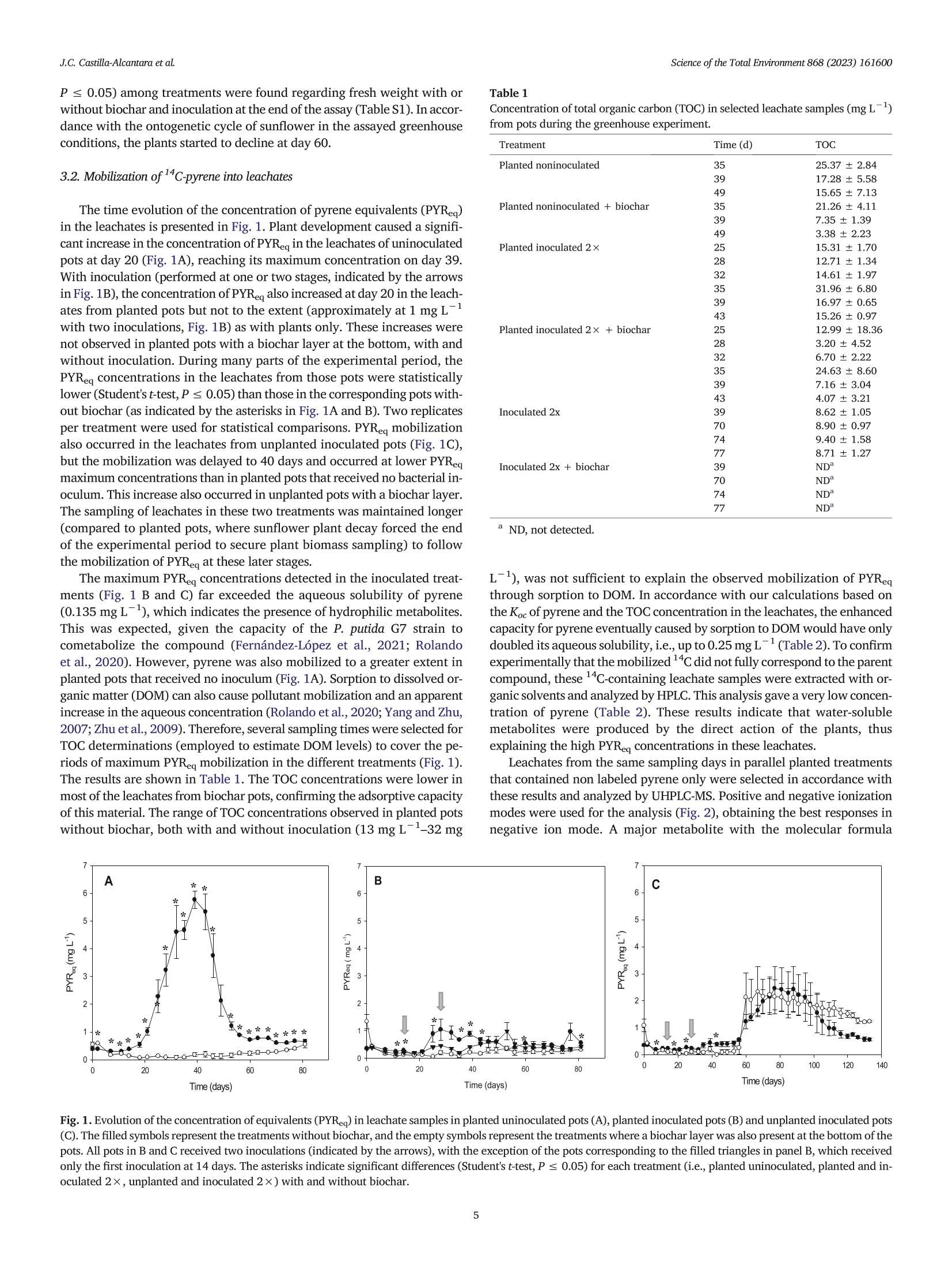
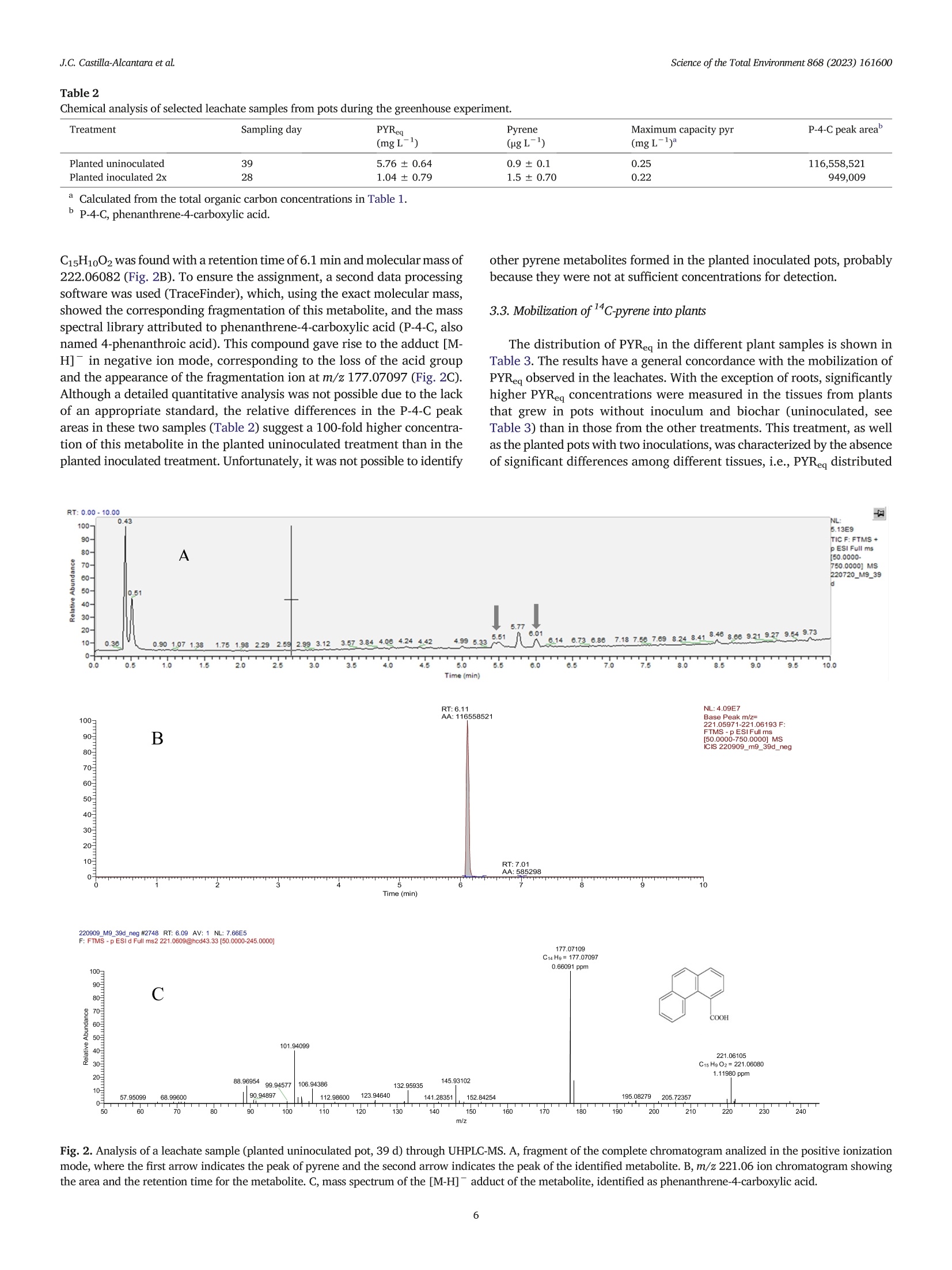
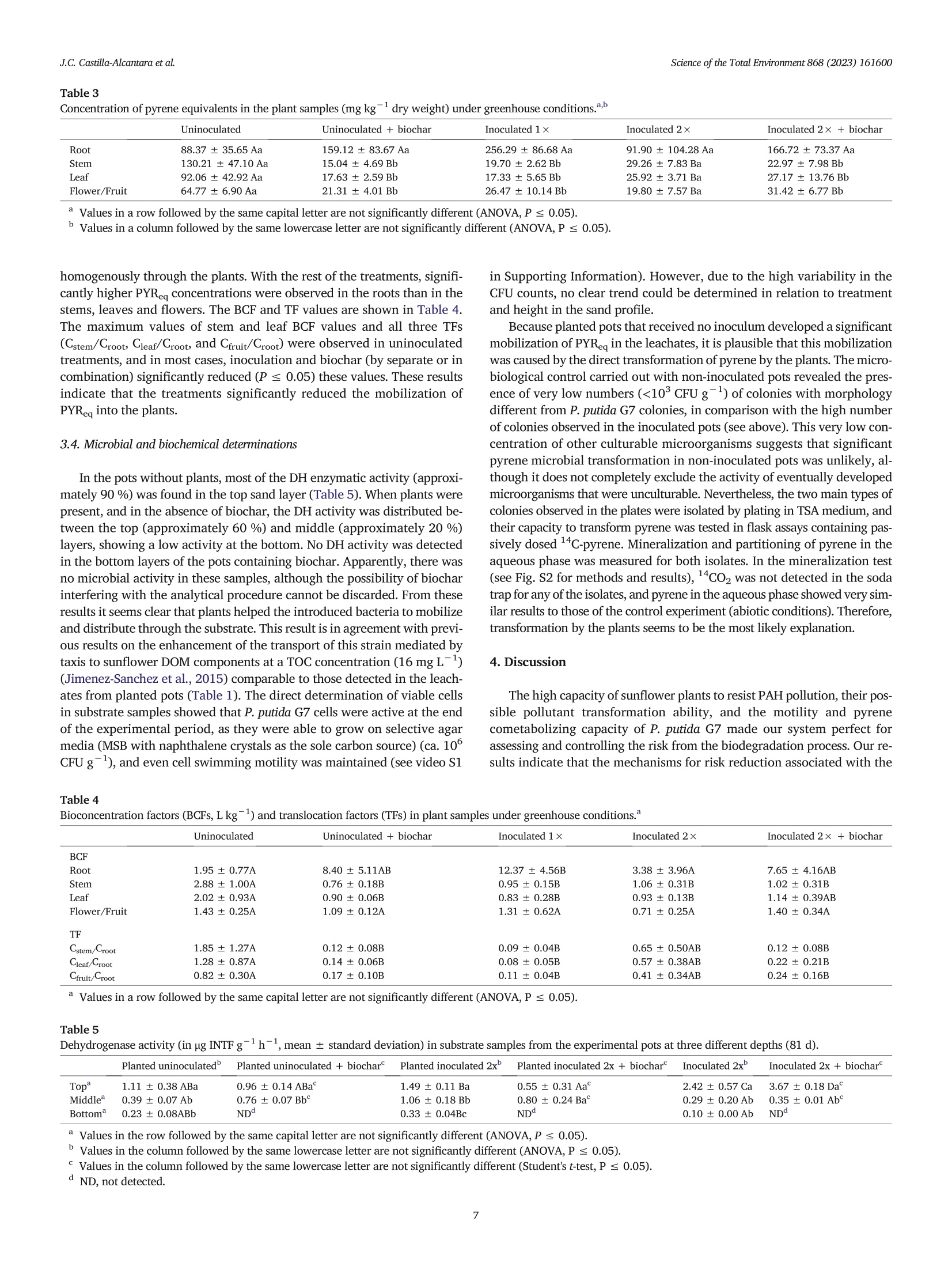
在植物-细菌生物炭系统模型中芘的生物转化和渗透过程中的风险降低Risk reductions during pyrene biotransformation and mobilization in a model plant-bacteria-biochar system采用格哈特公司 索克森Soxtherm 全自动快速固液萃取系统 基于固液萃取原理使用100 mL的丙酮/二氯甲烷(1:1v/v)测定5g选定渗滤液中12c-芘的残余浓度。The residual concentration of 12C-pyrene was also measured in selected leachate samples by exhaustive solid-liquid extraction using a subsample of 5 g and a mixture of 100 mL of acetone/dichloromethane (1:1 v/v) with a Soxtherm extractor (C. Gerhardt GmbH & Co. KG, Königswinter,Germany).Science of the Total Environment 868 (2023)161600 J.C. Castillla-Alcantara et alScience of the Total Environment 868 (2023)161600 Contents lists available at ScienceDirect Science of the Total Environment ELSEVIER j ournal homepage: www.elsevier.com/locate/scitotenv 在植物-细菌生物炭系统模型中芘的生物转化和渗透过程中的风险降低 Risk reductions during pyrene biotransformation and mobilization in amodel plant-bacteria-biochar system Jose Carlos Castilla-Alcantara , Rosa Posada-Baquero, Maria Balseiro-Romero, Carmen Fernandez-LópezJose Luis Garcia, Alicia Fernandez-Vazquez, John R. Parsons , Manuel Cantos, Jose Julio Ortega-Calvoa,* AInstituto de Recursos Naturales y Agrobiologia de Sevilla (IRNAS-CSIC), Seville, Spain. ”Centro Universitario de la Defensa, Universidad Politecnica de Cartagena, Santiago de la Ribera, Murcia, SpainInstitute for Biodiversity and Ecosystem Dynamics, University of Amsterdam, Amsterdam, the Netherlands ·Sunflowers transform distantly located14C-pyrene in soil. ·Root-mediated bacterial mobilization fa-cilitates cometabolism of this PAH. ·Pyrene metabolites appeared in leachatesand in plant tissues. ·Biochar retained metabolites and reducedbiotransformation risks. ·This new scenario has implications for riskreduction during bioremediation. ARTICLE INFO Editor: Frederic Coulon Keywords: BiodegradationRisk Polycyclic aromatic hydrocarbonsBioremediationSunflower Bacteria ABSTRACT The productive application of motile microorganisms for degrading hydrophobic contaminants in soil is one of themost promising processes in modern remediation due to its sustainability and low cost. However, the incomplete bio-degradation of the contaminants and the formation of the intermediary metabolites in the process may increase thetoxicity in soil during bioremediation, and motile inoculants may mobilize the pollutants through biosorption. There-fore, controlling these factors should be a fundamental part of soil remediation approaches. The aim of this study wasto evaluate the sources of risk associated with the cometabolism-based transformation of C-labeled pyrene by inoc-ulated Pseudomonas putida G7 and identify ways to minimize risk. Our model scenario examined the increase in bioac-cessibility to a distant source of contamination facilitated by sunflower (Helianthus annuus L.) roots. A biochar trap formobilized pollutant metabolites and bacteria has also been employed. The experimental design consisted of pots filledwith a layer of sand with C-labeled pyrene (88 mg kg) as a contamination focus located several centimeters fromthe inoculation point. Half of the pots included a biochar layer at the bottom. The pots were incubated in a greenhousewith sunflower plants and P. putida G7 bacteria. Pots with sunflower plants showed a higher biodegradation of pyrene,its mobilization as metabolites through the percolate and the roots, and bacterial mobilization toward the source ofcontamination, also resulting in increased pyrene transformation. In addition, the biochar layer efficiently reducedthe concentrations of pyrene metabolites collected in the leachates. Therefore, the combination of plants, motile bac-teria and biochar safely reduced the risk caused by the biological transformation of pyrene. * Corresponding author. E-mail address: jjortega@irnase.csic.es (J.J. Ortega-Calvo). http://dx.doi.org/10.1016/j.scitotenv.2023.161600 Received 21 November 2022; Received in revised form 23 December 2022; Accepted 10 January 2023 0048-9697/O 2023 The Authors. Published by Elsevier B.V. This is an open access article under the CC BY license (http://creativecommons.org/licenses/by/4.0/). Polycyclic aromatic hydrocarbons (PAHs) are one of the most commongroups of contaminants found in the environment. These chemical com-pounds are very persistent in soils due to their high hydrophobicity andhigh structural stability (Lawal, 2017; Ma et al., 2022). In this context,the bioavailability of these pollutants is an important subject in soil remedi-ation studies (Ortega-Calvo et al., 2013; Ortega-Calvo et al.,2015). Al-though the concept of bioavailability is often associated with techniquesto increase the solubilization of contaminants, it can also be increased bypromoting the dispersal of microorganisms through contaminated soil. Inthis process, bacterial chemotaxis plays an important role in the mobiliza-tion of PAH-degrading bacteria and the biodegradation of these contami-nants (Ahmad et al., 2020; Ortega-Calvo et al., 2003). Research has shown that many Pseudomonas species (Harwood et al.,1990; Sampedro et al., 2015), among others, exhibit chemotactic responsesto PAHs such as pyrene, anthracene or phenanthrene (Ortega-Calvo et al.,2003). Chemotaxis enhances the accessibility of the low-permeability re-gions of contaminated soils to bacteria, which may not be accessible by dis-persion alone and may increase the residence times of the bacteria in thecontamination sites and therefore, biodegradation (Lacal, 2017; Wanget al., 2016). However, the process of dispersing microorganisms in soilmay be quite complicated (Harms and Wick, 2006). Chemotaxis is essentialin bacterial transport in roots and is established as one of the major vectorsto promote bacterial dispersal (Dupuy and Silk, 2016). Kamilova et al.(Kamilova et al., 2005) showed that with the motile bacteria Pseudomonasfluorescens, Pseudomonas putida and Aeromonas hydrophila, tomato root col-onization was higher than that of the control with a nonmotile strain(Kamilova et al.,2005). A number of investigations have revealed the im-portance of bacterial taxis and their interactions with plant root exudatesand how they may improve the degradation efficiency (Neal et al., 2012;Sun et al., 2012; Zhang et al., 2013). Exudates include amino acids, organicacids and sugars, and some of them, such as organic acids, may modify thephysicochemical properties of the soil and enhance the attraction to thechemotactic bacteria to the pollutant focus (Hartmann et al., 2009). Rootexudates can also increase the degradation of PAHs in different ways: de-sorption of contaminants in soils, increasing bioavailability, can be en-hanced by the production of carboxylates; transformation of pollutants byenzymes secreted by plants and stimulation of the activity of microbialcommunities via C and N secreted by plants (Gkorezis et al., 2016). In this study, the cometabolic transformation of pyrene carried out by amotile microbial inoculant was taken as an example to assess risk that mayarise from the bioremediation of PAH-polluted soils. The application of acometabolic-competent inoculum based on motile bacteria may constitutean advantage in bioremediation, especially for high-molecular-weight(HMW) PAHs that are only degraded through cometabolism (Fernandez-Lopez et al., 2021). However, such biological processing may cause prob-lems if toxic and mobile metabolites are formed, thus constituting an addi-tional source of risk, different from the parent pollutant. For example, Tianet al. (2017) identified different pyrene products derived from the bacterialmetabolic activity of the pollutant as toxic to humans. Many of these metab-olites produced during the bioremediation process can even lead to changesin soil ecosystems, reducing the diversity of microbial organisms or decreasing seed production and root length in plants (Wang et al., 2021).Mo-tile bacteria can also mobilize pollutants through biosorption (Rolandoet al., 2020), increasing toxicity and the risk of contamination in less acces-sible areas such as groundwater (Kumar et al., 2022). We examined, in amodel scenario, how this risk could be reduced through an appropriate ar-rangement of plants and sorbent materials. In this paper, sunflower(Helianthus annuus L.) was used due to its strong tolerance to PAHs(Fernandez-Lopez et al., 2021; Ortega-Calvo et al., 2013; Ortega-Calvoet al., 2017). Some studies have reported significant phytoremediation efficiencies of sunflower species in petroleum-contaminated soil (Diab, 2008;Dickson et al., 2020; Sivaram et al., 2018). In addition to its rapid germination and long root system that increases the surface area for bacterial colo-nization (Maliszewska-Kordybach and Smreczak, 2000), the strong chemotactic response of P. putida G7 to sunflower root exudates has beenreported (Castilla-Alcantara et al., 2022). Therefore, sunflower and itshigher leaf evaporation level have been demonstrated to be adequate forthe experimental model system (Fernandez-Lopez et al., 2021; Tejeda-Agredano et al., 2013). In addition to the combined use of plants and bac-teria, biochar amendment has been established as another reference strat-egy for the remediation of PAH-contaminated soils and sediments (Biancoet al., 2021; Kong et al., 2018). Due to its eco-friendly use and low cost com-pared to physical and chemical techniques (Liu et al., 2011; Wu et al.,2017), biochar can be established as an added value to the bioremediationprocess. When adding biochar, an amount of metabolites could be absorbedonto the biochar, leading to a significant reduction in soil toxicity and bio-degradation enhancement (Kumar et al., 2022). We employed sunflower and the chemotactic bacterium P. putida G7 ina new model plant-bacteria-biochar system to understand the potentialsources of risk caused by motile inoculants and identify possible methodsfor risk minimization. This bacterial strain has been well studied in our lab-oratory under this context for chemotaxis to different substances, such assunflower plant exudates (Castilla-Alcantara et al., 2022; Jimenez-Sanchez et al., 2018), and its ability to degrade naphthalene and tocometabolize pyrene (Rolando et al., 2020). These characteristics makethis bacterium suitable for the purpose of our study. Therefore, we specifi-cally tested whether pyrene polar metabolites were produced in this systemunder greenhouse conditions and to what extent the potential risk frompyrene biodegradation could be reduced by incorporating biochar as a se-questering agent. Our experimental approach included, on the one hand,sand as a model substrate for plant growth and bacterial inoculation to min-imize the eventual interactions of the pollutant, its metabolites and the in-oculated bacteria with the substrate. On the other hand, 14c-tracertechniques were employed to facilitate the study of the transfer of pollutantcarbon to the substrate, leachates and plant tissues. 2. Materials and methods 2.1. Chemicals [4,5,9,10-4c]-pyrene (58.8 mCimmol-, radiochemical purity>98%)was purchased from Campro Scientific GmbH (Veenendaal, TheNetherlands), and c-pyrene (98% purity) was purchased from Sigma Al-drich (Madrid, Spain). Analytical-grade dichloromethane, acetonitrile,methanol and acetone were supplied by Fischer Chemical (Madrid,Spain). Washed sand was obtained from Scharlab, S.A. (Barcelona, Spain)and had the following characteristics: pH, 5.8; grain size, 300-350 um;and specific density, 2.66 g cm-3. Commercial biochar produced throughpyrolysis of birch, beech or oak wood was obtained from EGoS GmbH(Bot-trop, Germany). The material had a carbon content higher than 90 %and aninner surface area of 500 mg. The total PAH content was 0.2 mg kg,the humidity was 48.3 %, and the pH was 10. 2.2. Cultivation of bacteria The flagellated bacterium P. putida G7, which is able to use naphthaleneas the sole carbon and energy source, and degrade pyrene by cometabolism,was cultivated in MSB medium supplemented with salicylate and preparedfor greenhouse experiments as described previously (Fernandez-Lopezet al., 2021), with some modifications. Briefly, cultures were grown inthree stages to obtain a final volume of 7 L. Three bacterial cultures of100 mL were initially produced and transferred to three Erlenmeyer flasks(1 L) with 400 mL of MSB medium and then incubated for 24 h. The cul-tures were divided into seven 200-mL portions that were then transferredinto individual 2 L flasks, diluted with 800 mL of medium, and incubatedfor another 24 h to reach the early stationary phase (optical density at600 nm or OD600 of 0.5 or 5 × 10° cell mL-1). Cells were then harvestedby centrifugation at 5000 rpm for 10 min. The pellet was resuspended in100 mL of modified MSB (per liter: 670 mg of Na2HPO412H20,340 mgof KH2PO4, 80 mg of CaCl2, 33 mg of NHNO3, 112 mg of MgSO47H20, 5 mg of ZnSO47H20, 2.5 mg of Na2MoO42H20,0.22 mg of FeClg) withoutsalicylate to give a cell density of 5.58 ×10cell mL-1. This culture medium was modified to provide the necessary N requirement for plantgrowth and development, and still allowed a stable cell motility inP. putida, which was confirmed by light microscopy. Inoculation of 18 ofthe 26 total pots of the experiment with these suspensions (seeSection 2.3, greenhouse experiment) resulted in a cell concentration of1.43 ×10°cellkgsoil. 2.3. Greenhouse experiment 2.3.1. Experimental design The experiment was carried out in a greenhouse at 23±1°℃ using atotal of 26 pots. The pots were 18.5 cm high, and the maximum and mini-mum diameters were 21.5 and 15.0 cm, respectively. A hole at the bottomof each pot allowed for the collection of leachates during the experiment.Each pot received 4.8 kg of sand, which had a height of 11 cm upon pack-ing. The pots were classified into two different groups: those that receivedbiochar as a layer at the bottom and those without biochar. A diagram de-scribing the experiment is shown in SI (Fig. S1). In each group, a subgroupwas prepared with pots of sand loaded with C-andc-pyrene (14labeledpots, numbers 1-8 to 15-20) and another with only C-pyrene (12 unla-beled pots, numbers 9-14 to 21-26). The latter subgroup was used for esti-mations not requiring 4C-labeling (i.e., soil dehydrogenase (DH) activity,total organic carbon (TOC) measurements, metabolite analysis, and freshweight of plants). In each subgroup, there were three different treatments,each one performed in duplicate: planted inoculated pots (pots 1-4; 9-10;15-16;21-22), planted uninoculated pots (5-6; 11-12;17-18;23-24)and unplanted inoculated pots (7-8; 13-14;19-20;25-26). Neither thesand, the biochar, the percolation medium or water were sterilised as thepots were maintained in the greenhouse under non-sterile conditions. Be-cause the focus of this study was to monitor the transformation of 14cpyrene, no control pots were run without the contaminant. Nevertheless,preliminary germination and growth tests on sand with and without pyreneshowed no apparent effects of the chemical on the plants at the initial con-centration that was used in the greenhouse experiment, either as seed ger-mination, plant development or flowering. Before pot packing, pyrene was added to 500 g portions of sand thatwould later be introduced in each individual pot as a contaminated sandlayer. For this preparation, 1.5 mL of an acetone solution containing360,000 dpm of c-pyrene and sufficient unlabeled pyrene to give a con-centration 88 mg kg- in the contaminated layer of every pot, was dis-pensed onto a 10 g sand subsample. When the solvent was completelyevaporated, this subsample was mixed thoroughly with 40 g of sand in aglass jar. Then, this portion was homogenized with the rest of the sand por-tion (450 g) and shaken manually for 10 min to ensure homogenization. Inthe case of unlabeled pots, the spiking solution had only 12c-pyrene. Thecontaminated sand was introduced into the pots as a layer at 7 cm fromthe upper sand surface during packing. At the beginning of the experiment, the water content in each pot wasadjusted to 100 % water holding capacity with 1.2 L of Milli-Q water, dueto the rapid percolation capacity of the pure sand. The inoculation ofP. putida G7 was performed 14 days after of the introduction of the sun-flower seeds into the pots, once there was important root development.Then, 100 mL of a modified MSB suspension (described in Section 2.2) ofP. putida G7 cells was added to each pot. To maintain the moisture, 3 irriga-tions per week were necessary: 2 times per week with modified MSB me-dium for plant nutrition and bacterial maintenance (to coincide with thecollection of leachate) and once per week Milli-Q water was added to main-tain the humidity in the pots. A second inoculation with P. putida G7 wasperformed 28 days after seed introduction. The sunflower seeds were immersed in tap water and stirred for 30 minto eliminate anti- microorganism protection, and immediately, the coatswere manually removed. Fifteen of these naked seeds were used perplanted pot. Once the germination period was over, seven to thirteen plantsgrew per pot. Throughout the sunflower development cycle, the percentage of seed germination and the blooming evolution and stem length of plantswere separately evaluated for each treatment. 2.3.2. Sample collection Leachate sampling was performed to determine the 14c-activity at thebeginning, after 2 days, and then 2 days per week until the end of the exper-imental period. Leachates were collected by gravity immediately after eachirrigation event. Samples of sand and, if applicable, plants were collected atthe end of the experimental period (81 days for both planted and unplantedpots and 133 days for unplanted pots). Five sand samples were extractedwith a hand auger from each pot. These samples were then divided intothree subsamples and homogenized: top, corresponding to the first 2.5 cmof sand, middle, representing the next 1.5 cm; and bottom, representing re-maining sand to 11 cm. In the case of the pots with biochar, the third sam-ple was composed of a mixture of sand and biochar at the bottom of thepots. In unplanted pots, for which the experiment continued after the firstsampling, the holes in the pots were refilled with clean sand and markedto avoid additional sampling at those spots. Both the sand and leachate sam-ples were stored at -20 ℃ until analysis to prevent microbial activity.From each treatment, two complete plants per pot in the case of labeledpots were harvested at the end of the experiment; roots, stems, leaves,flowers and fruits were carefully separated for the oxidizer analysis; andfresh weight of each organ was obtained. All the plants that grew in the un-labeled treatment pots were also harvested at the end of the experiment,and their leaves, stems, roots and fruits were washed with distilled waterto calculate the fresh weight of each organ. An experimental control was run after the second inoculation of the pots(28 days) to establish possible bacterial percolation through irrigation. De-termination of the number of colony-forming units (CFUs) of P. putida G7 inleachate samples was carried out for planted and unplanted pots with orwithout biochar. A fraction of the leached liquid of the following two sam-ple collections after inoculation (4 and 7 days) from pots without c wascollected, inoculated on TSA agar plates, and incubated for 48 h at 30℃.The results showed that the maximum bacterial recovery in the leachateswas at day 4 in planted pots without biochar (2.81±0.66×10CFUmL-) in comparison with unplanted pots without biochar (9.45 ±4.66x10CFU mL-1). The presence of the biochar resulted in higher bacterialretention in both cases (1.37 ±0.27×105CFUmL-1 and 1.92±0.88×10 CFU mL-1 for planted and unplanted pots, respectively). However,many of the bacterial cells were retained in the pots because of the highcell density of the inoculum used. 2.3.3. Extraction and analysis of pyrene To measure the concentration of 4c-pyrene equivalents (PYReq) in theleachates, an aliquot (10 mL) was mixed with 10 mL of liquid scintillationcocktail (Ultima Gold XR, PerkinElmer). Radioactivity was measured by liq-uid scintillation (Beckman Instruments, Inc., Fullerton, Calif.; model LS6500 TD). In accordance to the specific activity of c-pyrene in the pots,1 dpm corresponded to 1 × 10-4mg PYRea. To determine 14C in sandand sunflower plant samples, a subsample weighing 1 g in the case ofsand samples or 0.2 g in the case of plant samples was combusted in an ox-idizer (307 Sample Oxidizer, Perkin Elmer, combustion for 5 min with O2).The samples obtained after combustion were measured by liquid scintilla-tion in a QUANTULUS 1220 (PerkinElmer) and for these samples the liquidscintillation cocktail used was Permafluor E+ from PerkinElmer. Prelimi-nary determinations with C-spiked sunflower plant samples resulted ina recovery of 100 % of the added radioactivity. The maximum capacity for pyrene of the leachates, assuming instanta-neous equilibration to reach the solubility of pyrene in water and to sorb tothe DOC components, was calculated as explained previously (Rolandoet al., 2020). Briefly, the predicted fraction of pyrene freely dissolved in theaqueous phase of bacterial suspensions at equilibrium (f) was calculated as where [DOC] is the concentration (in kg L-1) of dissolved organic carbon(DOC) determined experimentally in the leachates with a TOC-VC SH withan ASI-V auto sampler by Shimadzu Corporation (Tokyo, Japan) after filtra-tion through Whatman filter with 0.45 um pore diameter purchased fromSigma Aldrich (Madrid, Spain). The detection limit of this method was0.05 mg L-1 TOC. Koc is the solid-water distribution coefficient in Lkg-1.The log Koc of pyrene used for this equation was 4.7. The capacity for pyrenewas calculated as the ratio between the solubility in water of pyrene and f 2.3.4. Analysis of pyrene metabolites The analysis of leachates was performed with ultrahigh-performance liquid chromatography coupled to mass spectrometry (UHPLC-MS). The equip-ment setup included a Dionex Ultimate RS binary pump coupled to aquadrupole-Orbitrap QExactive hybrid mass spectrometer (Thermo FisherScientific, USA) equipped with an electrospray source (HESI-II) operating inpositive mode at 3.5 KV and in negative mode at -3.2 KV. UHPLC analysiswas carried out with a reversed-phase analytical column (Acquity UPLCBEH C18 (2.1 ×100 mm) with a 1.7 um particle size. The flow rate usedwas 0.4 mL min-. Chromatographic separation was performed using a bi-nary gradient of water (A) and methanol (B), both containing 0.15 % formicacid (v/v) to promote ionization. The elution profile consisted of 5 %B 1 min,then a linear gradient for 7.5 min until 100 % B was reached. The gradientwas maintained for 2 min, and finally, it was returned to the initial conditionsand maintained for 0.5 min. The chromatographic column was maintained at40℃ throughout the analysis. The injection volume was 5 pL. The sampleswere previously microfiltered with 0.2 um nylon filters. The acquisitionmethod in the QExactive mass spectrometer was performed using a data-dependent acquisition method (TOP5). Data files acquired by UHPLC wereprocessed by Compound Discoverer 3.2 software to identify unknown metab-olites and TraceFinder 5.1 software (Thermo Scientific) to determine com-pounds using molecular formulas and exact masses. 2.3.5. Calculation of the bioconcentration and translocation factors The transfer and distribution of organic chemicals in soil-plant systemshave been adequately explained by using bioconcentration factors (BCFs)and translocation factors (TFs). These factors are defined as the ratio ofthe chemical concentrations between plant biomass and soil solution andbetween leaves, stems and fruit biomass and root biomass, respectively(Doucette et al., 2018; Fernandez-Lopez et al., 2021; Gonzalez Garciaet al., 2018; Lin et al., 2006; Tao et al., 2008; Yang and Zhu, 2007). We hy-pothesized that any significant differences between treatments in the plantuptake or transfer rates of pyrene were indications of plant uptake of polametabolites generated by the modified bacterial accessibility andcometabolism of this PAH in soil. Therefore, the BCF for pyrene was calcu-lated based on “C-pyrene concentrations (see Section 2.3.3) and using thefollowing previously reported equation (Fernandez-Lopez et al., 2021;Gonzalez Garcia et al., 2019) with some modifications: where C is the concentration of pyrene equivalents (PYReq) in dry (rootsstems, leaves and fruits) plant biomass and Cs is the concentration ofPYReg (g L-l soil solution) calculated using the following equation: where A, is the amount of PYReg (mg) in the pots measured at the start ofthe experiment, A is the amount of PYReq (mg) in the leachate, V is the vol-ume of irrigation water (mL), V is the volume of leachate (mL), and A is theaverage amount of PYReg (mg) at the end of the experiment and was calcu-lated taking into account the specific weights in each part of the pots usingthe following equation: where A is the concentration of PYReq(mg kg) in the top of the pot mul-tiplied by the layer thickness in the top, A. is the concentration of PYReg(mg kg) in the center of the pot multiplied by the layer thickness in thecenter, At is the concentration of PYReg(mgkg) in the bottom of thepot multiplied by the layer thickness in the bottom, and W is the totalweight of sand in each pot (kg). The translocation factor (TF) was calculated to evaluate the transfer ofthe mass of pyrene from roots to leaves, stems and fruits, where a valueequal to or lower than 1 is optimal for phytostabilization, which involvesthe reduction in the mobility of PYReg in soil. The TF was calculated asthe ratio between the mass of pyrene in stems, leaves and fruits and thatin roots: where Cplant is the mass of PYReq(mg) per kg of dry stem, leaf and fruit bio-mass and C, is the mass of PYReq(mg) per kg of dry root biomass. 2.3.6. Microbial and biochemical determinations The number of P. putida cells was determined in sand samples from un-labeled pots at the end of the greenhouse experiment. Subsamples of sandcores at different heights (1g) were vortexed mixed with 3 mL of MSB me-dium, and agitated for 30 min at 150 rpm. After a proper time for sand de-cantation, serial dilutions of the supernatant were plated in MSB agar plateswith sterile naphthalene crystals on the lid. Bacterial motility was con-firmed using an AxioVert A1 phase contrast inverted microscope (Zeiss,Germany), and several videos were recorded using an Axiocam 305 color(Zeiss, Germany) (interface, Zen software blue edition). To exclude any pos-sible interference in the transformation of pyrene by cross-contamination,samples of non-inoculated pots (pots 11 and 12, Fig. S1) were plated onnaphthalene-containing solid medium with substrate samples obtained atthe end of the greenhouse experiment. Two main types of colonies differentto P. putida G7 were identified. This was expected because of the lack of ste-rility under greenhouse conditions. Nevertheless, the two main types of colo-nies observed in the plates were isolated and their capacity to transformpyrene was tested by radiorespirometry and partitioning in a passive dosingsystem (Rolando et al., 2020) in 100 mL flask assays containing 25 mL ofMSB and passively dosed C-pyrene (100,000 dpm) with a silicone O-ring.One milliliter samples were taken at different time periods from the sodatrap and the suspension, and mixed with 5 mL of scintillation liquid to deter-mine radioactivity as described above (see Section 2.3.3) The DH activity ofthe sand in the pots was determined by the reduction of the tetrazolium saltiodonitrotetrazolium chloride (INT) to yield iodonitrotetrazolium formazan(INTF) as described elsewhere (Garcia et al., 1994; Trevors, 1984) 3. Results P ≤ 0.05) among treatments were found regarding fresh weight with orwithout biochar and inoculation at the end of the assay (Table S1). In accordance with the ontogenetic cycle of sunflower in the assayed greenhouseconditions, the plants started to decline at day 60. 3.2. Mobilization of ““C-pyrene into leachates The time evolution of the concentration of pyrene equivalents (PYReq)in the leachates is presented in Fig. 1. Plant development caused a signifi-cant increase in the concentration of PYReg in the leachates of uninoculatedpots at day 20 (Fig. 1A), reaching its maximum concentration on day 39.With inoculation (performed at one or two stages, indicated by the arrowsin Fig. 1B), the concentration of PYReq also increased at day 20 in the leach-a-1tes from planted pots but not to the extent (approximately at1 mgLwith two inoculations, Fig. 1B) as with plants only. These increases werenot observed in planted pots with a biochar layer at the bottom, with andwithout inoculation. During many parts of the experimental period, thePYReq concentrations in the leachates from those pots were statisticallylower (Student's t-test, P≤0.05) than those in the corresponding pots with-out biochar (as indicated by the asterisks in Fig. 1A and B). Two replicatesper treatment were used for statistical comparisons. PYReq mobilizationalso occurred in the leachates from unplanted inoculated pots (Fig. 1C),but the mobilization was delayed to 40 days and occurred at lower PYReqmaximum concentrations than in planted pots that received no bacterial in-oculum. This increase also occurred in unplanted pots with a biochar layer.The sampling of leachates in these two treatments was maintained longer(compared to planted pots, where sunflower plant decay forced the endof the experimental period to secure plant biomass sampling) to followthe mobilization of PYReq at these later stages. YBThe maximum PYReq concentrations detected in the inoculated treat-ments (Fig. 1 B and C) far exceeded the aqueous solubility of pyrene(0.135 mg L ), which indicates the presence of hydrophilic metabolites.This was expected, given the capacity of the P. putida G7 strain tocometabolize the compound (Fernandez-López et al., 2021; Rolandoet al., 2020). However, pyrene was also mobilized to a greater extent inplanted pots that received no inoculum (Fig. 1A). Sorption to dissolved or-ganic matter (DOM) can also cause pollutant mobilization and an apparentincrease in the aqueous concentration (Rolando et al., 2020; Yang and Zhu,2007; Zhu et al., 2009). Therefore, several sampling times were selected forTOC determinations (employed to estimate DOM levels) to cover the pe-riods of maximum PYReq mobilization in the different treatments (Fig. 1).The results are shown in Table 1. The TOC concentrations were lower inmost of the leachates from biochar pots, confirming the adsorptive capacityof this material. The range of TOC concentrations observed in planted potswithout biochar, both with and without inoculation (13 mg L--32mg Table 1 Concentration of total organic carbon (TOC) in selected leachate samples (mg L-)from pots during the greenhouse experiment. Treatment Time (d) TOC Planted noninoculated 35 25.37±2.84 39 17.28±5.58 49 15.65±7.13 Planted noninoculated +biochar 35 21.26±4.11 39 7.35±1.39 49 3.38±2.23 Planted inoculated 2× 25 15.31±1.70 28 12.71±1.34 32 14.61±1.97 35 31.96±6.80 39 16.97±0.65 43 15.26±0.97 Planted inoculated 2x +biochar 25 12.99±18.36 28 3.20±4.52 32 6.70±2.22 35 24.63±8.60 39 7.16±3.04 43 4.07±3.21 Inoculated 2x 39 8.62±1.05 70 8.90±0.97 74 9.40±1.58 77 8.71±1.27 Inoculated 2x + biochar 39 ND 70 ND 74 ND 77 ND ND, not detected. L-), was not sufficient to explain the observed mobilization of PYRegthrough sorption to DOM. In accordance with our calculations based onthe Koc of pyrene and the TOC concentration in the leachates, the enhancedcapacity for pyrene eventually caused by sorption to DOM would have onlydoubled its aqueous solubility, i.e., up to 0.25 mg L-(Table 2). To confirmexperimentally that the mobilized 14c did not fully correspond to the parentcompound, these “C-containing leachate samples were extracted with or-ganic solvents and analyzed by HPLC. This analysis gave a very low concen-tration of pyrene (Table 2). These results indicate that water-solublemetabolites were produced by the direct action of the plants, thusexplaining the high PYReq concentrations in these leachates. Leachates from the same sampling days in parallel planted treatmentsthat contained non labeled pyrene only were selected in accordance withthese results and analyzed by UHPLC-MS. Positive and negative ionizationmodes were used for the analysis (Fig.2), obtaining the best responses innegative ion mode. A major metabolite with the molecular formula 5 4 3 2 Fig. 1. Evolution of the concentration of equivalents (PYReq) in leachate samples in planted uninoculated pots (A), planted inoculated pots (B) and unplanted inoculated pots(C). The filled symbols represent the treatments without biochar, and the empty symbols represent the treatments where a biochar layer was also present at the bottom of thepots. All pots in B and C received two inoculations (indicated by the arrows), with the exception of the pots corresponding to the filled triangles in panel B, which receivedonly the first inoculation at 14 days. The asterisks indicate significant differences (Student's t-test, P ≤ 0.05) for each treatment (i.e., planted uninoculated, planted and in-oculated 2×, unplanted and inoculated 2×) with and without biochar. Table 2 Chemical analysis of selected leachate samples from pots during the greenhouse experiment. Treatment Sampling day PYReq Pyrene Maximum capacity pyr P-4-C peak area” (mgL-) (rgL-) (mgL-)a Planted uninoculated 39 5.76±0.64 0.9±0.1 0.25 116,558,521 Planted inoculated 2x 28 1.04±0.79 1.5±0.70 0.22 949,009 aCalculated from the total organic carbon concentrations in Table 1. C15H1002 was found with a retention time of 6.1 min and molecular mass of222.06082 (Fig. 2B). To ensure the assignment, a second data processingsoftware was used (TraceFinder), which, using the exact molecular mass,showed the corresponding fragmentation of this metabolite, and the massspectral library attributed to phenanthrene-4-carboxylic acid (P-4-C, alsonamed 4-phenanthroic acid). This compound gave rise to the adduct [M-H]- in negative ion mode, corresponding to the loss of the acid groupand the appearance of the fragmentation ion at m/z 177.07097 (Fig. 2C).Although a detailed quantitative analysis was not possible due to the lackof an appropriate standard, the relative differences in the P-4-C peakareas in these two samples (Table 2) suggest a 100-fold higher concentra-tion of this metabolite in the planted uninoculated treatment than in theplanted inoculated treatment. Unfortunately, it was not possible to identify other pyrene metabolites formed in the planted inoculated pots, probablybecause they were not at sufficient concentrations for detection. 3.3. Mobilization of “C-pyrene into plants The distribution of PYReq in the different plant samples is shown inTable 3. The results have a general concordance with the mobilization ofPYReq observed in the leachates. With the exception of roots, significantlyhigher PYReq concentrations were measured in the tissues from plantsthat grew in pots without inoculum and biochar (uninoculated, seeTable 3) than in those from the other treatments. This treatment, as wellas the planted pots with two inoculations, was characterized by the absenceof significant differences among different tissues, i.e., PYReq distributed Fig. 2. Analysis of a leachate sample (planted uninoculated pot, 39 d) through UHPLC-MS. A, fragment of the complete chromatogram analized in the positive ionizationmode, where the first arrow indicates the peak of pyrene and the second arrow indicates the peak of the identified metabolite. B, m/z 221.06 ion chromatogram showingthe area and the retention time for the metabolite. C, mass spectrum of the [M-H]- adduct of the metabolite, identified as phenanthrene-4-carboxylic acid. Table 3Concentration of pyrene equivalents in the plant samples (mg kgdry weight) under greenhouse conditions.a.b Uninoculated Uninoculated + biochar Inoculated 1x Inoculated 2× Inoculated 2x + biochar Root 88.37±35.65 Aa 159.12±83.67 Aa 256.29±86.68 Aa 91.90±104.28 Aa 166.72±73.37 Aa Stem 130.21±47.10 Aa 15.04±4.69 Bb 19.70±2.62 Bb 29.26±7.83 Ba 22.97±7.98 Bb Leaf 92.06±42.92 Aa 17.63±2.59 Bb 17.33±5.65 Bb 25.92±3.71 Ba 27.17±13.76 Bb Flower/Fruit 64.77±6.90 Aa 21.31±4.01 Bb 26.47±10.14 Bb 19.80±7.57 Ba 31.42 ±6.77 Bb Values in a row followed by the same capital letter are not significantly different (ANOVA, P≤0.05). Values in a column followed by the same lowercase letter are not significantly different (ANOVA,P≤ 0.05). homogenously through the plants. With the rest of the treatments, signifi-cantly higher PYReq concentrations were observed in the roots than in thestems, leaves and flowers. The BCF and TF values are shown in Table 4.The maximum values of stem and leaf BCF values and all three TFs(Cstem/Croot Gleaf/Croot, and Cfruit/Croot) were observed in uninoculatedtreatments, and in most cases, inoculation and biochar (by separate or incombination) significantly reduced (P ≤ 0.05) these values. These resultsindicate that the treatments significantly reduced the mobilization ofPYReq into the plants. 3.4. Microbial and biochemical determinations In the pots without plants, most of the DH enzymatic activity (approxi-mately 90 %) was found in the top sand layer (Table 5). When plants werepresent, and in the absence of biochar, the DH activity was distributed be-tween the top (approximately 60 %) and middle (approximately 20 %)layers, showing a low activity at the bottom. No DH activity was detectedin the bottom layers of the pots containing biochar. Apparently, there wasno microbial activity in these samples, although the possibility of biocharinterfering with the analytical procedure cannot be discarded. From theseresults it seems clear that plants helped the introduced bacteria to mobilizeand distribute through the substrate. This result is in agreement with previ-ous results on the enhancement of the transport of this strain mediated bytaxis to sunflower DOM components at a TOC concentration (16mgL)(Jimenez-Sanchez et al.,2015) comparable to those detected in the leach-ates from planted pots (Table 1). The direct determination of viable cellsin substrate samples showed that P. putida G7 cells were active at the endof the experimental period, as they were able to grow on selective agarmedia (MSB with naphthalene crystals as the sole carbon source) (ca. 10° CFU g ), and even cell swimming motility was maintained (see video S1 in Supporting Information). However, due to the high variability in theCFU counts, no clear trend could be determined in relation to treatmentand height in the sand profile. Because planted pots that received no inoculum developed a significantmobilization of PYReq in the leachates, it is plausible that this mobilizationwas caused by the direct transformation of pyrene by the plants. The micro-biological control carried out with non-inoculated pots revealed the pres-ence of very low numbers (<10°CFU g) of colonies with morphologydifferent from P. putida G7 colonies, in comparison with the high numberof colonies observed in the inoculated pots (see above). This very low con-centration of other culturable microorganisms suggests that significantpyrene microbial transformation in non-inoculated pots was unlikely, al-though it does not completely exclude the activity of eventually developedmicroorganisms that were unculturable. Nevertheless, the two main types ofcolonies observed in the plates were isolated by plating in TSA medium, andtheir capacity to transform pyrene was tested in flask assays containing pas-sively dosed c-pyrene. Mineralization and partitioning of pyrene in theaqueous phase was measured for both isolates. In the mineralization test(see Fig. S2 for methods and results),4Co2 was not detected in the sodatrap for any of the isolates, and pyrene in the aqueous phase showed very sim-ilar results to those of the control experiment (abiotic conditions). Therefore,transformation by the plants seems to be the most likely explanation. 4. Discussion The high capacity of sunflower plants to resist PAH pollution, their pos-sible pollutant transformation ability, and the motility and pyrenecometabolizing capacity of P. putida G7 made our system perfect forassessing and controlling the risk from the biodegradation process. Our re-sults indicate that the mechanisms for risk reduction associated with the Table 4 Bioconcentration factors (BCFs, Lkg) and translocation factors (TFs) in plant samples under greenhouse conditions.a Uninoculated Uninoculated + biochar Inoculated 1× Inoculated 2× Inoculated2x + biochar BCF Root 1.95±0.77A 8.40±5.11AB 12.37±4.56B 3.38±3.96A 7.65±4.16AB Stem 2.88±1.00A 0.76±0.18B 0.95±0.15B 1.06±0.31B 1.02±0.31B Leaf 2.02±0.93A 0.90±0.06B 0.83±0.28B 0.93 ±0.13B 1.14±0.39AB Flower/Fruit 1.43±0.25A 1.09±0.12A 1.31±0.62A 0.71±0.25A 1.40±0.34A TF Cstem/Croot 1.85±1.27A 0.12±0.08B 0.09±0.04B 0.65±0.50AB 0.12±0.08B Cleaf/Croot 1.28±0.87A 0.14±0.06B 0.08±0.05B 0.57±0.38AB 0.22±0.21B Cfruit/Croot 0.82±0.30A 0.17±0.10B 0.11±0.04B 0.41±0.34AB 0.24±0.16B aValues in a row followed by the same capital letter are not significantly different (ANOVA, P ≤0.05). Table 5 Dehydrogenase activity (in pg INTFg h,mean ± standard deviation) in substrate samples from the experimental pots at three different depths (81 d). Planted uninoculated Planted uninoculated + biochar Planted inoculated 2x² Planted inoculated 2x + biochar Inoculated 2x” Inoculated 2x + biochar° Top 1.11±0.38 ABa 0.96 ±0.14 ABa° 1.49±0.11 Ba 0.55±0.31 Aa 2.42±0.57 Ca 3.67±0.18 Da Middle 0.39± 0.07 Ab 0.76± 0.07 Bb° 1.06±0.18 Bb 0.80 ± 0.24 Ba 0.29 ±0.20 Ab 0.35±0.01 Ab Bottom° 0.23±0.08ABb ND 0.33 ±0.04Bc ND 0.10 ±0.00 Ab ND Values in the row followed by the same capital letter are not significantly different (ANOVA, P≤ 0.05). Values in the column followed by the same lowercase letter are not significantly different (ANOVA, P≤ 0.05). Values in the column followed by the same lowercase letter are not significantly different (Student's t-test, P≤ 0.05). leaching of pyrene metabolites resided, on the one hand, on plantmicroorganism interactions and, on the other hand, on the trapping capac-ity of the biochar. The presence of sunflower roots also facilitated microbialdispersal to the source of contamination, probably due to the chemotacticability of the P. putida G7 strain to many of the components of the exudatesproduced by the plant (Fernandez-López et al., 2021; Jimenez-Sanchezet al., 2015). Also, irrigation may have physically affected the dispersal inplanted pots, particularly as the plant roots grew downward toward thepyrene-sand layer. Taking into account the hazardous nature of many ofthe substances produced during the pollutant degradation process, the ad-dition of biochar as a sequestering agent reduces the risk associated withthe decontamination process. In this way, the novel system proposed inthis study may open up new possibilities for the future control of risk asso-ciated with bioremediation in contaminated soils. The enhanced concentra-tion of PYReq observed in the leachates from planted pots after inoculation(Fig. 1B) may reflect the cometabolic activity of P. putida G7 cells once theyreached the distant pyrene source, which was facilitated by the plants. Thispromoting effect of sunflower plants on the transport and pyrene-cometabolic activity of P. putida G7 cells in soil has already been observed(Fernandez-López et al., 2021). Our results extend those findings by show-ing a significant reduction in PYReq concentrations in the leachates fromplanted, inoculated pots equipped with a biochar layer, therefore indicatingthat the mobilized metabolites were efficiently retained in the pots. The enhancement observed under greenhouse conditions at later stagesin inoculated pots that had no plants can be explained by the activity of the bacteria that were also transported, still to a limited extent, to the contam-inated zone. Such an increase was also observed in inoculated potsequipped with the biochar layer, which can be explained by the fact thatthis material was only stable and therefore retained its sorption capacityduring the first two months of experimentation. This may not only be an en-vironmentally sustainable option due to the biodegradability of the prod-uct, avoiding the persistence of exogenous components in the soil for longtimes, it could also provide an ecological niche that allows for controlledbacterial deposition at the source of contamination. In this way, the highavailability of the nutrients provided by the biochar can promote the micro-bial activity of the bacterial cells retained in the product, increasing thedegrading capacity of the pollutants present in the material. However, themicrobial and biochemical determinations performed at the end of the ex-perimental period indicate that, without plants, most of the bacterial inoc-ulant was retained in the upper layers of the pots. The limited transport ofthis strain through porous materials and the enhanced mobilization and ac-tivity as a result of tactic reactions have been examined in a variety of sce-narios, including sand columns (Jimenez-Sanchez et al., 2015; Rolandoet al., 2020) and sunflower-planted soil (Fernandez-López et al., 2021).Our results extend those findings by showing that mobilized bacteria andmetabolites can efficiently be controlled through a combination of plantsand adsorbent phases. 5. Conclusions The present proof-of-concept study shows a new integrated model forbioremediation where the role of bacterial motility combined with sun-flower activity generates a synergic response for pyrene degradation. Wedemonstrated that the action of the plant and the bacteria was effective inthe degradation of pyrene as well as that the plant facilitated the dispersalof the motile bacteria. In addition, the use of a biochar-based sequesteringtrap for the metabolites produced during biodegradation made it possibleto reduce the leaching risk associated with the decontamination process.These results provide new insights into the bioremediation of PAH-polluted matrices, where biologically caused environmental risk is consid-ered fundamental in the process. The feasibility of this greenhouse experi-ment under real field conditions is made effective due to the low cost ofthe materials used in the process, providing new tools for future applicationin the remediation sector. Our results suggest improved nature-based solu-tions for the treatment of soils polluted by PAHs and other hydrophobic pol-lutants and show that the use of plant-bacteria-sorbent arrangements canconstitute a valid alternative to control the risk of such contamination.The results could also be applied in wastewater reuse for soil irrigationwhere the biological transformation of organic contaminants introducedin soil may cause further environmental and human risk. Supplementary data to this article can be found online at https://doi.org/10.1016/j.scitotenv.2023.161600. CRediT authorship contribution statement Jose Carlos Castilla-Alcantara: Conceptualization, Methodology,Data curation, Writing-original draft. Rosa Posada-Baquero: Conceptu-alization, Methodology, Data curation, Writing -original draft. MariaBalseiro-Romero: Conceptualization, Methodology, Data curation.Carmen Fernandez-Lopez: Conceptualization, Writing-original draft.Jose Luis Garcia: Methodology, Data curation, Writing - original draft.Alicia Fernandez-Vazquez: Methodology, Data curation. John R. Par-sons: Conceptualization, Writing-original draft. Manuel Cantos: Concep-tualization, Methodology, Data curation, Writing -original draft. JoseJulio Ortega-Calvo: Writing-review & editing, Funding acquisition. Data availability No data was used for the research described in the article. Declaration of competing interest The authors declare that they have no known competing financial inter-ests or personal relationships that could have appeared to influence thework reported in this paper. Acknowledgments We would like to thank the Spanish Ministries of Economy and Compet-itiveness (CGL2016-77497-R) and Science and Innovation (PID2019-109700RB-C21 and PRIMA project PCI2020-111967) for supporting thiswork. We also acknowledge the European Union's Horizon 2020 researchand innovation program (Marie Sklodowska-Curie grant agreement no.895340; BIOTAC project). References Ahmad, F., Zhu, D., Sun, J.,2020. Bacterial chemotaxis: a way forward to aromatic com-pounds biodegradation. Environ. Sci. Eur. 32 (1), 52. Alagic, s.C., Maluckov, B.S., Radojicic, V.B., 2015. How can plants manage polycyclic aro-matic hydrocarbons? May these effects represent a useful tool for an effective soil reme-diation? A review. Clean Techn. Environ. Policy 17 (3), 597-614. Al-Shaikh, A.J., Jamal, M.T., 2020. Bioaugmentation of halophilic consortia for the degrada-tion of petroleum hydrocarbons and petroleum wastewater treatment. Int. J. Adv. Res.Biol. Sci. 7 (9),97-112. Balseiro-Romero, M., Gkorezis, P., Kidd, P.S., Van Hamme, J., Weyens, N., Monterroso, C.,Vangronsveld, J., 2017. Use of plant growth promoting bacterial strains to improveCytisus striatus and Lupinus luteus development for potential application inphytoremediation. Sci. Total Environ. 581-582, 676-688. Bianco, F., Race, M., Papirio, S., Oleszczuk, P., Esposito, G., 2021. The addition of biochar as asustainable strategy for the remediation of PAH-contaminated sediments. Chemosphere263,128274. Castilla-Alcantara, J.C., Akbari, A., Ghoshal, S., Ortega-Calvo, J.J., 2022. Role of tactic re-sponse on the mobilization of motile bacteria through micrometer-sized pores. Sci.Total Environ. 832,154938. Diab, E.A., 2008. Phytoremediation of oil contaminated desert soil using the rhizosphere ef-fects. Glob.J.Environ.Res. 2 (2),66-73. Dickson, U.J., Coffey, M., George Mortimer, R.J., Smith, B., Ray, N., Di Bonito, M.,2020. Investigating the potential of sunflower species, fermented palm wine andPleurotus ostreatus for treatment of petroleum-contaminated soil. Chemosphere240,124881. Doucette, W.J., Shunthirasingham, C., Dettenmaier, E.M., Zaleski, R.T., Fantke, P., Arnot, J.A.,2018. A review of measured bioaccumulation data on terrestrial plants for organicchemicals: metrics, variability, and the need for standardized measurement protocols. En-viron. Toxicol. Chem. 37 (1), 21-33. Dupuy, L.X., Silk, W.K., 2016. Mechanisms of early microbial establishment on growing rootsurfaces. Vadose Zone J. 15 (2),1-13. Fernandez-Lopez, C., Posada-Baquero, R., Garcia, J.L., Castilla-Alcantara, J.C., Cantos, M.,Ortega-Calvo, J.J., 2021. Root-mediated bacterial accessibility and cometabolism ofpyrene in soil. Sci. Total Environ. 760, 143408. Garcia, C., Hernandez, T., Costa,F., 1994. Microbial activity in soils under Mediterranean en-vironmental conditions. Soil Biol. Biochem. 26 (9),1185-1191. Gkorezis, P., Daghio, M., Franzetti, A., Van Hamme, J.D., Sillen, W., Vangronsveld, J., 2016.The interaction between plants and bacteria in the remediation of petroleum hydrocar-bons: an environmental perspective. Front. Microbiol. 7 1836-1836. Glick, B.R., 2012. Plant growth-promoting bacteria: mechanisms and applications. Scientifica2012,963401. Gonzalez Garcia, M., Fernandez-Lopez, C., Pedrero-Salcedo, F., Alarcon, J.J., 2018. Absorp-tion of carbamazepine and diclofenac in hydroponically cultivated lettuces and humanhealth risk assessment. Agric. Water Manag. 206,42-47. Gonzalez Garcia, M., Fernandez-Lopez, C., Polesel, F., Trapp, S., 2019. Predicting the uptakeof emerging organic contaminants in vegetables irrigated with treated wastewater-im-plications for food safety assessment. Environ. Res. 172, 175-181. Hadibarata, T., Kristanti, R.A., 2013. Biodegradation and metabolite transformation of pyrene by basidiomycetes fungal isolate Armillaria sp. F022. Bioprocess Biosyst. Eng. 36 (4),461-468.Haritash, A.K., Kaushik, C.P., 2009. Biodegradation aspects of polycyclic aromatic hydrocar-bons (PAHs): a review. J. Hazard. Mater. 169(1-3),1-15. Harms, H., Wick, L.Y., 2006. Dispersing pollutant-degrading bacteria in contaminated soilwithout touching it. Eng.Life Sci. 6 (3), 252-260. Hartmann, A., Schmid, M., Tuinen, D.v., Berg, G., 2009. Plant-driven selection of microbes.Plant Soil 321 (1),235-257. Harwood, C.S., Parales, R.E., Dispensa, M., 1990. Chemotaxis of Pseudomonas putida towardchlorinated benzoates. Appl. Environ.Microbiol. 56 (5), 1501-1503. Hiickelhoven, R., Schuphan, I., Thiede, B., Schmidt, B., 1997. Biotransformation of pyrene by cellcultures of soybean (Glycine max L.), wheat (Triticum aestivum L.), Jimsonweed (Datura stra-monium L.), and purple foxglove (Digitalis purpurea L.). J. Agric. Food Chem. 45 (1),263-269. Jimenez-Sanchez, C., Wick, L.Y., Cantos, M., Ortega-Calvo,J.-J., 2015. Impact of dissolved or-ganic matter on bacterial tactic motility, attachment, and transport. Environ.Sci.Technol.49 (7), 4498-4505. Jimenez-Sanchez, C., Wick, L.Y., Ortega-Calvo, J.J., 2018. Impact of chemoeffectors on bacte-rial motility, transport, and contaminant degradation in sand-filled percolation columns.Environ. Sci. Technol. 52, 10673-10679. Kamilova, F., Validov, S., Azarova, T., Mulders, I., Lugtenberg, B., 2005. Enrichment for en-hanced competitive plant root tip colonizers selects for a new class of biocontrol bacteria.Environ. Microbiol.7 (11),1809-1817. Kathi, S., Khan, A.B., 2011. Phytoremediation approaches to PAH contaminated soil. IndianJ. Sci. Technol. 4(1),56-63. Kong, L., Gao, Y., Zhou, Q., Zhao, X., Sun, Z., 2018. Biochar accelerates PAHs biodegradationin petroleum-polluted soil by biostimulation strategy. J. Hazard. Mater. 343,276-284. Kuhn, A., Ballach, H.J., Wittig, R., 2004. Studies in the biodegradation of 5 PAHs (phenan-threne, pyrene, fluoranthene, chrysene und benzo(a)pyrene) in the presence of rootedpoplar cuttings. Environ. Sci. Pollut. Res. Int. 11 (1), 22-32. Kumar,M.,Bolan,N., Jasemizad,T., Padhye, L.P., Sridharan, S., Singh, L., Bolan, S.,O'Connor,J., Zhao, H., Shaheen, S.M., Song, H., Siddique, K.H.M., Wang, H., Kirkham, M.B.,Rinklebe,J., 2022.Mobilization of contaminants: potential for soil remediation and unin-tended consequences. Sci. Total Environ. 839,156373. Lacal, J., 2017. The potential of hydrocarbon chemotaxis to increase bioavailability and bio-degradation efficiency. In: Krell, T. (Ed.), Cellular Ecophysiology of Microbe. Springer In-ternational Publishing, Cham, pp. 1-14. Lawal, A.T., 2017. Polycyclic aromatic hydrocarbons. A review. Cogent Environ. Sci. 3 (1),1339841. Lin, D., Zhu, L., He, W., Tu, Y., 2006. Tea plant uptake and translocation of polycyclic aro-matic hydrocarbons from water and around air. J. Agric. Food Chem. 54 (10),3658-3662. Liu, P.-W.G., Chang, T.C., Whang, L.-M., Kao, C.-H., Pan, P.-T., Cheng, S.-S., 2011. Bioremedi-ation of petroleum hydrocarbon contaminated soil: effects of strategies and microbialcommunity shift. Int. Biodeterior. Biodegradation 65 (8), 1119-1127. Ma, L., Yao, L., Li, Y., 2022. Bioremediation of a polycyclic aromatic hydrocarbon-contaminated urban soil: degradation dynamics and phytotransformation pathways.J. Soils Sediments 22 (3), 797-808. Maliszewska-Kordybach, B., Smreczak, B., 2000. Ecotoxicological activity of soils pollutedwith polycyclic aromatic hydrocarbons (PAHs)-effect on plants. Environ. Technol. 21(10),1099-1110. Nakajima, D., Kojima, E., Iwaya, S., Suzuki, J., Suzuki, S., 1996. Presence of 1-hydroxypyreneconjugates in woody plant leaves and seasonal changes in their concentrations. Environ.Sci.Technol. 30 (5),1675-1679. Neal, A.L., Ahmad,., Gordon-Weeks, R., Ton, J., 2012. Benzoxazinoids in root exudates ofmaize attract Pseudomonas putida to the rhizosphere. PLOS ONE 7 (4), e35498. Ortega-Calvo, J.J., Marchenko, A.I., Vorobyov, A.V., Borovick, R.V., 2003. Chemotaxis inpolycyclic aromatic hydrocarbon-degrading bacteria isolated from coal-tar- and oil-polluted rhizospheres. FEMS Microbiol. Ecol. 44 (3), 373-381. Ortega-Calvo, J.J., Tejeda-Agredano, M.C., Jimenez-Sanchez, C., Congiu, E.,Sungthong, R., Niqui-Arroyo, J.L., Cantos, M., 2013. Is it possible to increase bio-availability but not environmental risk of PAHs in bioremediation? J. Hazard.Mater. 261.733-745. Ortega-Calvo, J.J., Posada-Baquero, R., Garcia, J.L., Cantos, M., 2017. Bioavailability of poly-cyclic aromatic hydrocarbons in soil as affected by microorganisms and plants. In: Lukac,M., Grenni, P., Gamboni, M. (Eds.), Soil Biological Communities And Ecosystem Resil- ience. Springer International Publishing, Cham,pp. 305-319.Ortega-Calvo,J.-J., Harmsen, J., Parsons, J.R., Semple, K.T., Aitken, M.D., Ajao, C., Eadsforth, C., Galay-Burgos, M., Naidu,R., Oliver,R., Peijnenburg, W.J.G.M., Rombke, J., Streck, G.,Versonnen, B., 2015. From bioavailability science to regulation of organic chemicals. En-viron. Sci. Technol. 49, 10255-10264. Posada-Baquero, R., Jimenez-Volkerink, S.N., Garcia, J.L., Vila, J., Cantos, M., Grifoll, M.,Ortega-Calvo,J.J., 2020.Rhizosphere-enhanced biosurfactant action on slowly desorbingPAHs in contaminated soil. Sci. Total Environ. 720, 137608. Posada-Baquero, R., Semple, K.T., Ternero, M., Ortega-Calvo, J.J., 2022. Determining the bio-availability of benzo(a)pyrene through standardized desorption extraction in a certifiedreference contaminated soil. Sci. Total Environ. 803,150025. Rolando, L., Vila, J., Posada-Baquero, R., Castilla-Alcantara, J.C., Barra Caracciolo, A., Ortega-Calvo, J.-J., 2020. Impact of bacterial motility on biosorption and cometabolism ofpyrene in a porous medium. Sci. Total Environ. 717, 137210. Sampedro, I., Parales, R.E., Krell, T., Hill, J.E., 2015. Pseudomonas chemotaxis. FEMSMicrobiol. Rev. 39 (1), 17-46. Sivaram, A.K., Logeshwaran, P., Lockington, R., Naidu, R., Megharaj, M., 2018. Impact ofplant photosystems in the remediation of benzo[a]pyrene and pyrene spiked soils.Chemosphere 193, 625-634. Sun, S., Wang,J., Zhu, L., Liao, D., Gu, M., Ren, L., Kapulnik, Y., Xu, G., 2012. An active factorfrom tomato root exudates plays an important role in efficient establishment of mycorrhi-zal symbiosis. PLOS ONE 7 (8),e43385. Tao, Y., Zhang, S., Wang, Z., Christie, P., 2008. Predicting bioavailability of PAHs in soils towheat roots with triolein-embedded cellulose acetate membranes and comparison withchemical extraction. J. Agric. Food Chem. 56 (22),10817-10823. Tejeda-Agredano, M.C., Gallego, S., Vila, J., Grifoll, M., Ortega-Calvo, J.-J., Cantos, M., 2013.Influence of the sunflower rhizosphere on the biodegradation of PAHs in soil. Soil Biol.Biochem. 57, 830-840. Tian, Z., Gold, A., Nakamura, J.,Zhang, Z., Vila,J., Singleton, D.R., Collins, L.B., Aitken, M.D.,2017. Nontarget analysis reveals a bacterial metabolite of pyrene implicated in the geno-toxicity of contaminated soil after bioremediation. Environ.Sci.Technol. 51 (12),7091-7100. Trevors, J.T., 1984. Dehydrogenase activity in soil: a comparison between the INT and TTCassay. Soil Biol. Biochem. 16 (6), 673-674. Wang, X., Lanning, L.M., Ford, R.M., 2016. Enhanced retention of chemotactic bacteria in apore network with residual NAPL contamination. Environ. Sci. Technol. 50 (1),165-172. Zhang, N., Wang, D., Liu, Y., Li, S., Shen, Q., Zhang, R., 2013. Effects of different plantroot exudates and their organic acid components on chemotaxis, biofilm formationand colonization by beneficial rhizosphere-associated bacterial strains. Plant Soil374,689-700. Zhu, Y., Zhang, S., Huang, H., Wen, B., 2009. Effects of maize root exudates and organic acidsLCon the desorption of phenanthrene from soils. J. Environ. Sci. 21 (7), 920-926.
确定







还剩8页未读,是否继续阅读?

产品配置单



中国格哈特为您提供《土壤渗滤液中12C-芘含量的检测》,该方案主要用于土壤中有机污染物检测,参考标准《HJ 784-2016 土壤和沉积物 多环芳烃的测定 高效液相色谱法》,《土壤渗滤液中12C-芘含量的检测》用到的仪器有格哈特全自动快速溶剂萃取仪Sox416、德国加液器MM、滤纸筒
推荐专场
快速溶剂萃取设备/快速溶剂萃取仪
更多


相关方案
更多
该厂商其他方案
更多