








方案详情
文
非监督的MS/MS二级谱图的相似性代谢轮廓图分析快速找出了17种以前未曾分析出的代谢物,并发现了羟基肉桂酰胺在AtACT-AtdTX18超表达路径中的合成机制。
方案详情

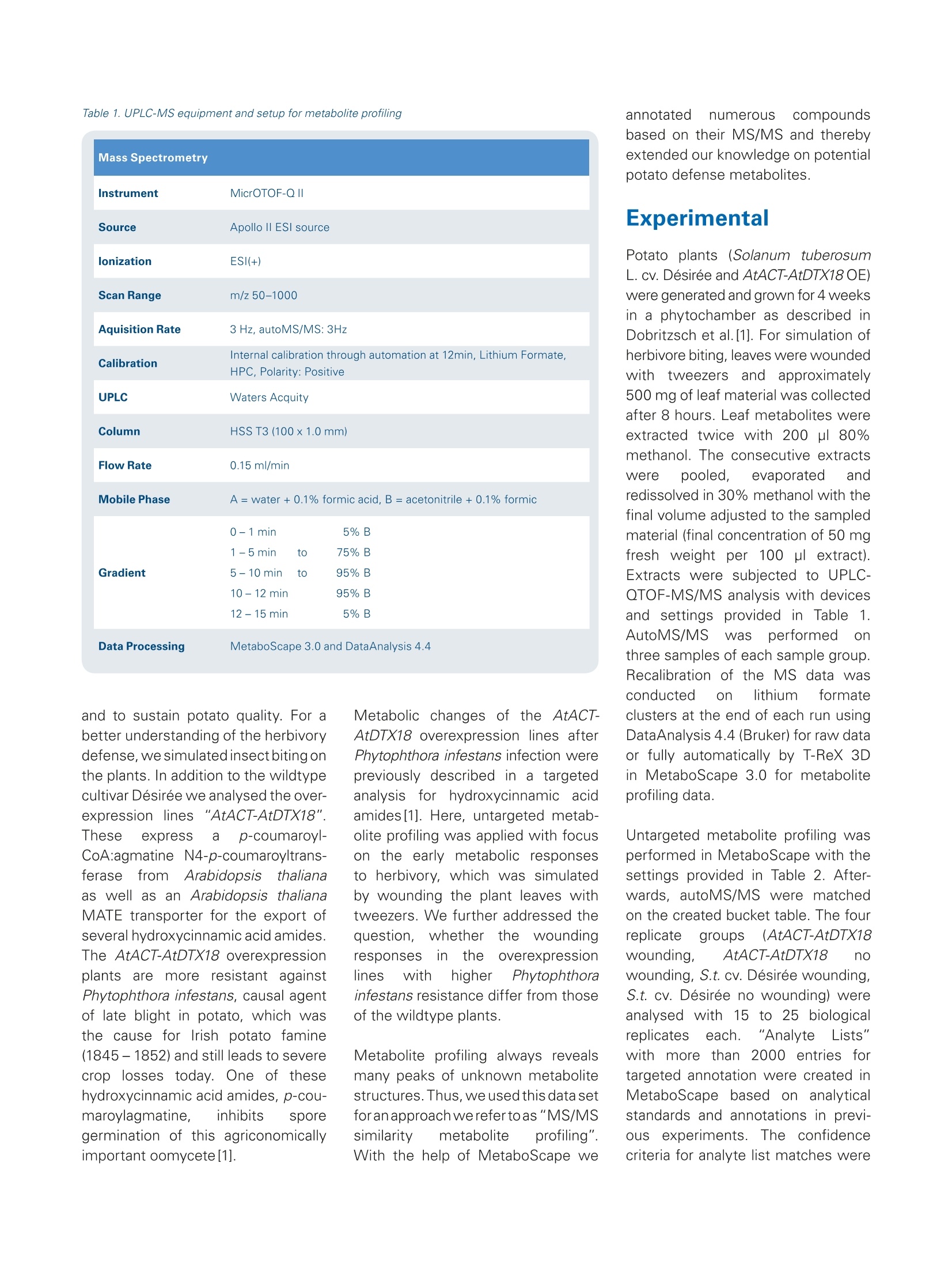
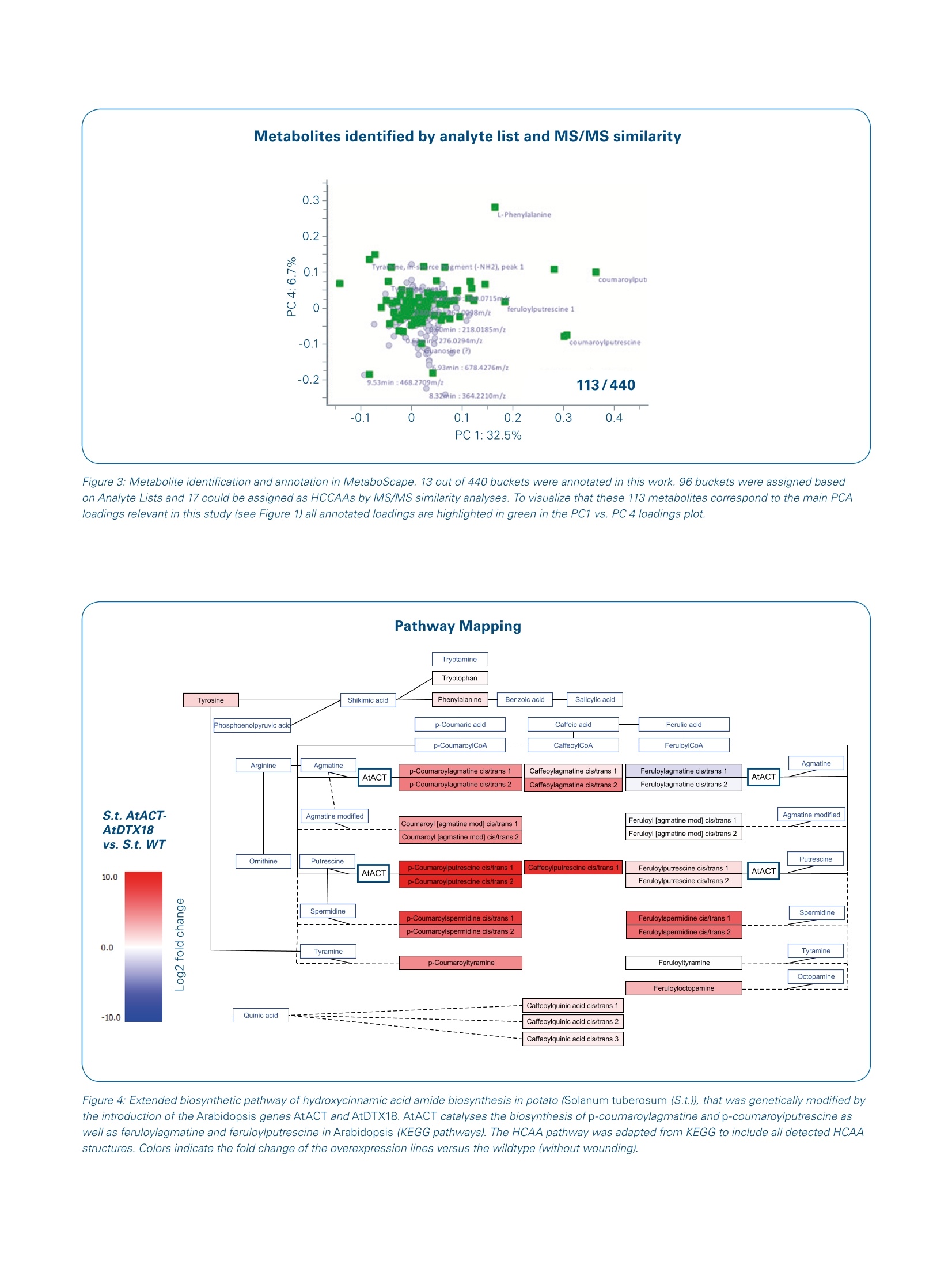
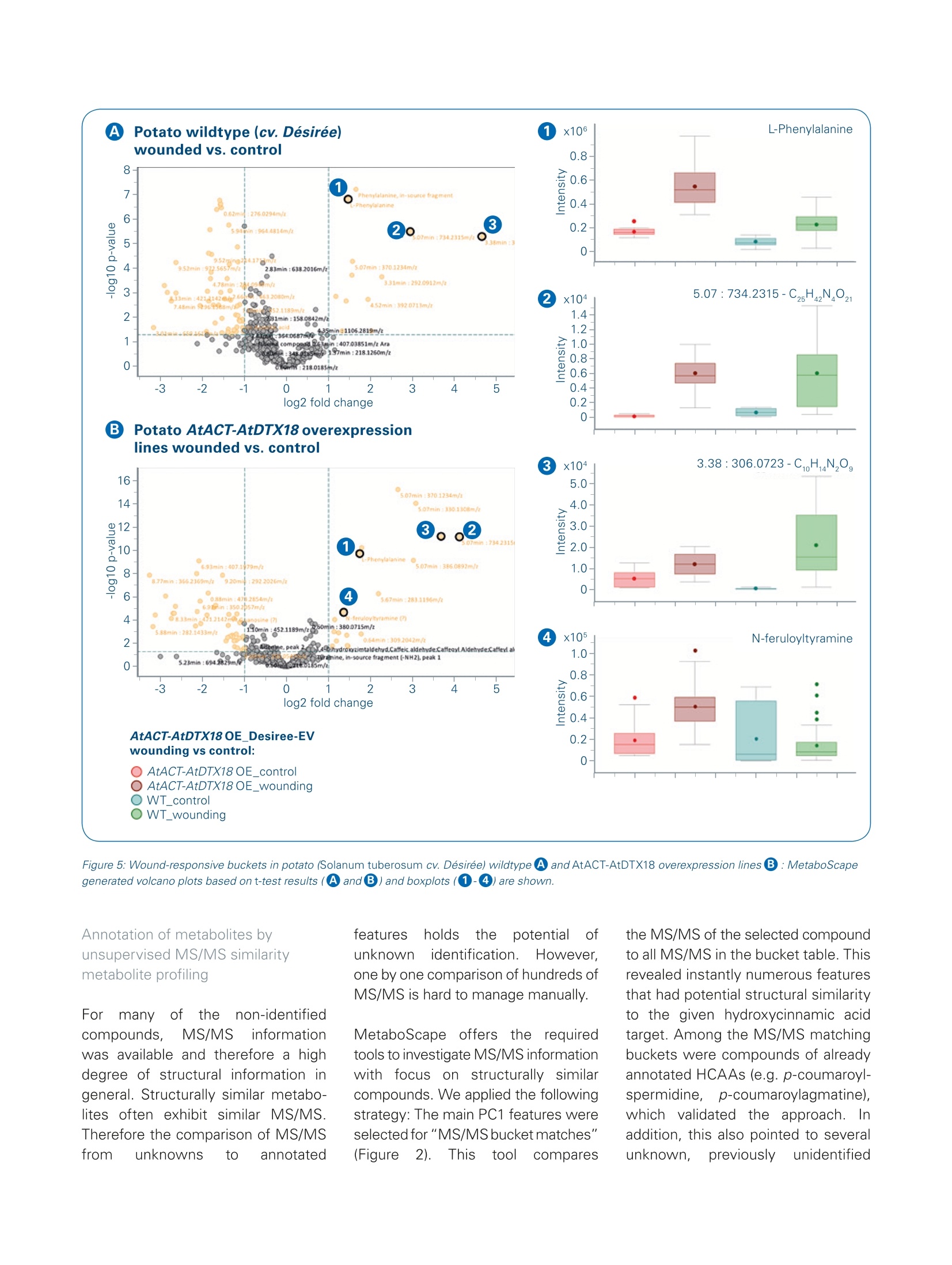
ms.sales.bdal@bruker.com-www.bruker.com How potato fights its enemies - UnsupervisedMS/MS similarity metabolite profiling revealsnovel defense-related metabolites in potato Solanum tuberosum (potato) wildtype and overexpression lines with an enhancedPhytophtora infestans resistance (1) were analysed for their metabolic response tosimulated herbivory. Introduction Using the data acquiredd withBruker LC-ESI-Q-TOF MS andMS/MS instrumentationaandmethods, the MetaboScapesoftware provided an efficientplatform for the straightforwardlinkage of structural similaritiesand metabolite abundances. The described data mining approachwe refer to as "unsupervised MS/MS similarity metabolite profil-ing" enabled a rapid annotation of17 metabolites which were pre-viously unknown in our analyses. Solanum tuberosum (potato) isanimportant crop for humannutrition with high agronomical andcultural importance. Resistanceagainst biotic attacks is desirableto reduce pesticide treatments Keywords.Potato metabolites,MS/MS similarities-metabolite similarities,untargeted metaboliteprofiling, MetaboScape,UPLC-QTOF-MS,microTOF-QI1.DataAnalysis ( A u t h o rs : D r . K a r i n G o r z o l ka, D r . S tef a n i e Do l l , Dr . S teff e n N e u man n , U l r i ke S m olk a , Pr o f .D r . S a b ine Ros ah l, P r o f . D r. D ie r k S c h e el; L e i b n i z- In s ti t u t e f o r P la nt B io ch emi st ry , S tres s a n d Dev e l op men t a l B i o l o g y , H a ll e , G e rm a n y . ) Mass Spectrometry Instrument MicrOTOF-QII Source Apollo Il ESI source lonization ESI(+) Scan Range m/z 50-1000 Aquisition Rate 3Hz, autoMS/MS: 3Hz Calibration Internal calibration through automation at 12min, Lithium Formate, HPC, Polarity: Positive UPLC Waters Acquity Column HSS T3 (100x1.0mm) Flow Rate 0.15 ml/min Mobile Phase A =water + 0.1% formic acid, B = acetonitrile +0.1% formic 0-1min 5%B 1-5 min to 75%B Gradient 5- 10 min to 95% B 10 -12 min 95% B 12-15 min 5%B Data Processing MetaboScape 3.0 and DataAnalysis 4.4 and to sustain potato quality. For abetter understanding of the herbivorydefense, we simulated insect biting onthe plants. In addition to the wildtypecultivar Desiree we analysed the over-expression lines:s"AtACT-AtDTX18".These express a p-coumaroyl-CoA:agmatine N4-p-coumaroyltrans-ferase from Arabidopsis thalianaas well as an Arabidopsis thalianaMATE transporter for the export ofseveral hydroxycinnamic acid amides.The AtACT-AtDTX18 overexpressionplants are more resistant againstPhytophthora infestans, causal agentof late blight in potato, which wasthe cause for Irish potato famine(1845-1852) and still leads to severecrop losses today. One of thesehydroxycinnamic acid amides, p-cou-maroylagmatine, inhibits sporegermination of this agriconomicallyimportant oomycete[1]. Metabolic changes of the AtACT-AtDTX18 overexpression lines afterPhytophthora infestans infection werepreviously described in a targetedanalysiss forrhydroxycinnamic acidamides [1]. Here, untargeted metab-olite profiling was applied with focuson the early metabolic responsesto herbivory, which was simulatedby wounding the plant leaves withtweezers. We further addressed thequestion,,whetherthe woundingresponses inttheoverexpressionlines with higher Phytophthorainfestans resistance differ from thoseof the wildtype plants Metabolite profiling always revealsmany peaks of unknown metabolitestructures. Thus, weusedthis data setforan approach werefer toas"MS/MSsimilarity metabolite profiling".With the help of MetaboScape we annotated numerouS compoundsbased on their MS/MS and therebyextended our knowledge on potentialpotato defense metabolites. Experimental Potato plants (Solanum tuberosumL. cv. Desiree and AtACT-AtDTX18OE)weregenerated and grown for 4 weeksin a phytochamber as described inDobritzsch et al.[1]. For simulation ofherbivore biting, leaves were woundedwithh tweezers and approximately500 mg of leaf material was collectedafter 8 hours. Leaf metabolites wereextracted twice with 200 pl 80%methanol. The consecutive extractsWerepooled, evaporated andredissolved in 30% methanol with thefinal volume adjusted to the sampledmaterial (final concentration of 50 mgfresh weight per 100 pl extract).Extracts were subjected to UPLC-QTOF-MS/MS analysis with devicesand settings provided innTable1.AutoMS/MS was performedonthree samples of each sample group.Recalibration of the MS data wasconducted on lithium tormateclusters at the end of each run usingDataAnalysis 4.4 (Bruker) for raw dataor fully automatically by T-ReX 3Din MetaboScape 3.0 for metaboliteprofiling data. Untargeted metabolite profiling wasperformed in MetaboScape with thesettings provided in Table 2. After-wards, autoMS/MS were matchedon the created bucket table. The fourreplicate groups (AtACT-AtDTX18wounding, AtACT-AtDTX18 nowounding, S.t. cv. Desiree wounding,S.t. cv. Desiree no wounding) wereanalysed with. 15 to 25 biologicalreplicates each. "AnalyteeLists"with more than 2000 entries fortargeted annotation were created inMetaboScape based on analyticalstandards and annotations in previ-ous experiments.The confidencecriteria for analyte list matches were (annotation quality: very good/accept-able): m/z: 2 ppm/20 ppm, retentiontime: (0.1min/0.2min, mSigma:20/60, MS/MS score: 900/800. Pathway maps wereecdrawn inPathVisio 3.3.0 [2] based on informationfrom KEGG pathways [3] (arginine andproline metabolism/phenylpropanoidbiosynthesis) with Solanum tuberosumand Arabidopsis thaliana as referenceorganisms. Results MetaboScape-Metabolite profilingof wounded and unwoundedSolanum tuberosum plants With the selected settings for peakpicking (Table 2) we focused on thehigher abundant leaf metabolites. TheMetaboScape T-ReX 3D peak pickingalgorithm generated a bucket tableof 440 region complete extractedfeatures. For most of these, the MS/MS were assigned from autoMS/MS.The analyte lists provided annotationsfor 96 of the 440 metabolites thatmatched the criteria of correct retentiontime, m/z and isotopic pattern. Theintuitive graphical annotation qualitysymbols enabled a fast and reliable Table 2. MetaboScape 3.0 processing parameters Intensity threshold [counts] 1500 Minimum peak length [spectra] 10 Minimum peak length (recursive) [spectra] 8 Minimum #Features for Extraction 31 Presence of features in minimum # of analyses 31 Lock mass calibration false Mass calibration true Calib. RT start 723 Calib.RTend 732 S.t. AtACT-AtDTX18 overexpression Filter by sample group vs S.t. cv. Desiree Minimal value of features in sample group (type) PERCENTAGE Minimal value of features in sample group 75 Area true Primary lon [M+H]+ Seedlons [M+Na]+ EIC correlation 0.9 Mass range: Start [m/z] 50 Mass range:End [m/z] 1000 Retention time range:Start [min] 0.6 Retention time range: End [min] 10 Perform MS/MS import false Group by collision energy false MS/MS import method maxsum Figure 2:MS/MS similarity metabolite profiling. The MS/MS of p-coumaroylputrescine (cis/trans-isomer 2) was selected for (MS/MS bucket matches》(bucket highlighted in blue in the center). Hydroxycinnamic acid amides (HCAAs) are stereoisomers and occur as two LC-separated peaks, which are termedas ucis/trans 1 or 2. Of the 11 matching MS/MS, 6 compounds were identified as HCAAs [compounds①.②.③[2 isomers]and 4)[2 isomers]].Compound ⑤ [2 isomers] could be described by in-depth MS/MS analysis as coumaroylagmatine with a modified agmatine residue; the compounds in box 6were flagged as tentative HCAA-like compounds. review of the identification confi-dence. Thus,1estraightforwardthedataaprocessingmnMetaboscapeprovided identification for approxi-mately 25% of the features. Thisemphasized thatitpotato contitainsmanyspecific metabolites whiichare not yet covered by the utilizedAnalyte List. A principal component analysis (PCA)provided a first impression of sample similarity and treatment-dependentmetabolicCdifferences. A cleardistinction between the wildtype andthe overexpression lines (OE) waspresent in PC 1, whereas in PC 4 thegroups of wounded vs. unwoundedplantsi separated(Figure1).Theloading values of the PCA depictedcoumaroylagmatine, coumaroyl-putrescine,feruloylputrescineandcaffeoylputrescineaas the mostrelevant PC 1 metabolites. The en- hancement of these hydroxycinnamicacid amides (HCAAs) in the OE lineswas described in a targeted analysisby our group before [1]. From thiswork we knew about the high MS/MSsimilarity of HCAAs with indicativemain fragments and systematic neutrallosses. This was the basis for a novelinvestigation of the data, which werefer to as"unsupervised MS/MSsimilarity metabolite profiling". Figure 3: Metabolite identification and annotation in MetaboScape. 13 out of 440 buckets were annotated in this work. 96 buckets were assigned basedon Analyte Lists and 17 could be assigned as HCCAAs by MS/MS similarity analyses. To visualize that these 113 metabolites correspond to the main PCAloadings relevant in this study (see Figure 1) all annotated loadings are highlighted in green in the PC1 vs. PC 4 loadings plot. Figure 4: Extended biosynthetic pathway of hydroxycinnamic acid amide biosynthesis in potato (Solanum tuberosum (S.t.)), that was genetically modified bythe introduction of the Arabidopsis genes AtACT and AtDTX18. AtACT catalyses the biosynthesis of p-coumaroylagmatine and p-coumaroylputrescine aswell as feruloylagmatine and feruloylputrescine in Arabidopsis (KEGG pathways). The HCAA pathway was adapted from KEGG to include all detected HCAAstructures. Colors indicate the fold change of the overexpression lines versus the wildtype (without wounding). Figure 5: Wound-responsive buckets in potato (Solanum tuberosum cv. Desiree) wildtype AandAtACT-AtDTX18 overexpression lines B:MetaboScapegenerated volcano plots based on t-test results (A and B)and boxplots (①-4) are shown. Annotation of metabolites byunsupervised MS/MS similaritymetabolite profiling' ForrrmanyoDf theennon-identifiedcompounds, MS/MS informationwas available and therefore a highdegree of structural information ingeneral. Structurally similar metabo-lites often exhibit similar MS/MS.Therefore the comparison of MS/MSfrom unknowns to annotated l teatures holdsSthe potentiaal ofunknownidentification.1.However.one by one comparison of hundreds ofMS/MS is hard to manage manually. MetaboScape offerss the requiredtools to investigate MS/MS informationwith focus on structurally similarcompounds. We applied the followingstrategy: The main PC1 features wereselected for"MS/MS bucket matches"(Figure 2). This tool compares the MS/MS of the selected compoundto all MS/MS in the bucket table. Thisrevealed instantly numerous featuresthat had potential structural similarityto the given hydroxycinnamic acidtarget. Among the MS/MS matchingbuckets were compounds of alreadyannotated HCAAs (e.g. p-coumaroyl-spermidine,, p-coumaroylagmatine),which validated the approach. Inaddition, this also pointed to severalunknown, Ppreviously unidentified compounds with MS/MS similarity.MetaboScape enables to flagcompounds of interest for examplefor detailed investigation at a laterstage. As shown in Figure 2 thisflagging was used for highlightingseveral HCAA-like compounds. Afteradditional detailed investigation ofthe MS/MS,,wecouldannotatenumerous so far undescribed struc-tures of the HCAA compound family.We applied this approach on otherteatures(feruloyl-l- cand caffeoyl-amides) and found in total 24 HCAAs.Consequently, tlthe MetaboScapesoftware allowed extracting manycompoundswith one Cclick thatprovided】 structuralsimilarity.Asshown in Figure 3intotal 113metabolites could be annotated. lt should be emphasized that nearlyall HCAAs were of higher abundanceIntheoverexpression lines. Weused the tool "Pathway Mapping" inMetaboScape to visualize tlthiscommon enhancement and toemphasize common biosyntheticorigins.A pathway map was extendedby tlthe many additional novelmetabolitesthatwereannotatedin course of the MS/MS similarityapproach described above. Fur-thermore,we couldiirntegrate thereactions that were catalyzed by theoverexpressed AtACT gene productaccording to information derived fromKEGG[3]. The fold change mappingin MetaboScape (Figure 4) pointedto two main findings: A) The AtACToverexpression did not only influencethe abundance of its reported directmetabolic products; nearly all HCAAswere changed and increased in the OElines. B) Feruloylagmatine, althoughdescribed to be a catalytic product ofthe AtACT enzyme, was decreasedafter overexpression of the enzyme.Thus. immediate downstreamdegradation of feruloylagmatinemight be present, precursors mightbe limited or the Arabidopsis enzymemight exhibit distinct substrate specificity in the metabolic contextof potato.This underlines, that theMetaboScapee workflowprovidednumerous findings for exciting followup experiments. Wound-induced metabolites inpotato The metabolic changes in responseto wounding became obviousbythe comparison of control and treat-ment in both genotypes (wildtypeand overexpression lines) separately.The t-test results revealed commonwound-induced features (Figure 5).Phenylalanine was significantlyincreased after wounding in both WTand OE plants (Figure 5, compound 1),whichwas6 also implied byv thePCA with phenylalanine as PC 4loading value (Figure 1). The molecularformulae of other common wound-induceddfeatures wereputativelyassigned with Smart Formula(Figure 5,compounds2and 33).Afew metabolites -wwereewound-induced in the AtACT-AtDTX18over-expression lines, but not significantly in the wildtype plants 8 hours afterwounding. Theemost interestingfeature was feruloyltyramine (Figure 5,compound 4). Feruloyltyramine is acompound of the HCAA metaboliteclade and was already described asa Phytophthora infestans-responsivemetabolite in potato by Keller et al.,1996[4] and as wound-responsivecompound in potato tubers by Negrelet al., 1993[5]. In contrast to bothgroups, we did not observe significantincreases of feruloyltyramine in thewounded WT plants, which might beexplained by an earlier sampling timepoint (8 hours instead of 24 and 48hours (Keller et al.), 6 days (Negrelet al.)) ordifferential rreactionstowards the oomycete elicitor and inresponse to wounding. Acknowledgement We thank Bruker Daltonics (Bremen),especially Dr. Aiko Barsch, Dr. NikolasKessler and Dr.Heiko Neuweger, fortheir helpful support and scientificdiscussion. Conclusion Using the data acquired with Bruker LC-ESI-Q-TOF MS and MS/MSinstrumentation and methods, the MetaboScape software providedan efficient platform for the straightforward linkage of structuralsimilarities and metabolite abundances. We used various statisticaltools (PCA, t-test) in MetaboScape to focus on relevant features.The unsupervised MS/MS similarity metabolite profiling approachenabled a rapid annotation of 17 metabolites which were previouslyunknown in our analyses. The common abundance patterns together with common MS/MSpatterns pointed to an overflow of the hydroxycinnamic acid amidebiosynthesis in the AtACT-AtDTX18 overexpression lines. This is inperfect accordance with the function of the overexpressed enzymeAtACT that is responsible for the biosynthesis of four HCAAs. Theincrease of the many other HCAAs might be a metabolic side effectof the plant to compensate for this overflow and to bypass toxiceffects for the cell. MetaboScape was very valuable for easy dataprocessing, the annotation of many formerly unknown compoundsand an intuitive visualization of metabolic changes. Learn More You are looking for further Information?Check out the link or scan the QR code for more details. www.bruker.com/metaboscape References [1] Dobritzsch M, Lubken T, Eschen-Lippold L, Gorzolka K, Blum E, Matern A, et al. MATE TransporterDependent Export of Hydroxycinnamic Acid Amides. Plant Cell. 2016;28(2):583-96. [2]Kutmon M, van lersel MP Bohler A, KelderT, Nunes N, Pico AR, et al. PathVisio 3: An Extendable Pathway Analysis Toolbox. PLOS Computational Biology. 2015;11(2):e1004085.[3]|Kanehisa M, Furumichi M, Tanabe M, Sato Y, Morishima K. KEGG: new perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Research.2017;45(D1):D353-D61.[4]Keller H, Hohlfeld H,WrayV, Hahlbrock K, Scheel D, Strack D. Changes in the accumulation of soluble and cell wall-bound phenolics in elicitor-treated cell suspension cultures and fungus-infected leaves of Solanum tuberosum. Phytochemistry. 1996;42(2):389-96.5 [5]Negrel J,Javelle F Paynot M.Wound-Induced Tyramine Hydroxycinnamoy/ Transferase in Potato(Solanum tuberosum) Tuber discs. Journal of Plant Physiology. 1993;142(5):518-24. Please contact your local representative for availability in your country.For Research Use Only. Not for Use in Clinical Diagnostic Procedures. Bruker Daltonik GmbH Bruker Scientific LLC Bremen · GermanyBillerica, MA·USAPhone +49 (0)421-2205-0Phone +1 (978) 663-3660 Solanum tuberosum (potato) wildtype and overexpression lines with an enhanced Phytophtora infestans resistance were analysed for their metabolic response to simulated herbivory.• Using the data acquired with Bruker LC-ESI-Q-TOF MS and MS/MS instrumentation and methods, the MetaboScape software provided an efficient platform for the straightforward linkage of structural similarities and metabolite abundances. We used various statistical tools (PCA, t-test) in MetaboScape to focus on relevant features. The unsupervised MS/MS similarity metabolite profiling approach enabled a rapid annotation of 17 metabolites which were previously unknown in our analyses.• The common abundance patterns together with common MS/MS patterns pointed to an overflow of the hydroxycinnamic acid amide biosynthesis in the AtACT-AtDTX18 overexpression lines. This is in perfect accordance with the function of the overexpressed enzyme AtACT that is responsible for the biosynthesis of four HCAAs. The increase of the many other HCAAs might be a metabolic side effect of the plant to compensate for this overflow and to bypass toxic effects for the cell. MetaboScape was very valuable for easy data processing, the annotation of many formerly unknown compounds and an intuitive visualization of metabolic changes.
确定





还剩6页未读,是否继续阅读?

布鲁克·道尔顿(Bruker Daltonics)为您提供《土豆中防御有关的代谢物检测方案(生物质谱)》,该方案主要用于蔬菜中防御有关的代谢物检测,参考标准--,《土豆中防御有关的代谢物检测方案(生物质谱)》用到的仪器有
相关方案
更多
该厂商其他方案
更多