








方案详情
文
本文译自Werner等科学家在美国化学学会会刊(ACS)上发表的一篇综述,比较详实系统介绍了超高分辨率显微技术在神经科学上的最新应用进展。我们在此文基础上进行了编译整理。因文章较长,我们将分期陆续介绍。本期介绍第一部分。
方案详情

本文译自Werner等科学家在美国化学学会会刊(ACS)上发表的一篇综述,比较详实系统介绍了超高分辨率显微技术在神经科学上的最新应用进展。我们在此文基础上进行了编译整理。因文章较长,我们将分期陆续介绍。本期介绍第二部分。《Super-resolving Microscopy in Neuroscience》荧光成像技术在我们理解神经系统中起着关键作用。各种超分辨率显微镜方法和专用荧光探针的出现使得能够以迄今为止无与伦比的分辨率直接洞察细胞亚区室中的神经元结构和蛋白质排列。神经元的超分辨率可视化技术揭示了对细胞骨架组成、分布、运动性和膜蛋白信号传导、突触下结构和功能以及神经元-胶质细胞相互作用的新认识。自身免疫性和神经退行性疾病模型中明确的分子靶点为使用新型和创新的成像方法深入研究疾病病理生理学提供了极好的起点。超分辨率显微镜在人脑样本和临床生物标志物检测中的应用仍处于起步阶段,但为神经学和神经科学的转化研究提供了新的机会。在这篇综述中,作者描述了超分辨显微镜在过去的二十年里是如何提高我们对神经元和大脑功能和功能障碍的理解的。2015年,Ed Boyden及其同事描述了另一种突破荧光显微镜分辨率限制的创新策略,称为膨胀样品显微镜成像技术(ExM)。ExM基于物理样品扩增,通过将蛋白质连接到带电聚丙烯酰胺凝胶中,然后使用蛋白酶进行轻度机械破坏并在水中扩增。最初的方法是将三功能连接物交联到凝胶基质上,将标记信息转移到凝胶上。所使用的三官能接头含有甲基丙烯酰基、荧光标记和用于与互补寡核苷酸杂交的寡核苷酸,所述互补寡核苷酸附着于用于蛋白质标记的抗体。将免疫染色的细胞或组织包埋在含有丙烯酸钠的单体溶液中(使凝胶具有高吸水性)后,加入共聚单体丙烯酰胺和交联剂N,N ′-亚甲基双(丙烯酰胺)。使用四甲基乙二胺(TEMED)作为催化剂和过硫酸铵(APS)用于聚合引发,在升高的温度下聚合这些单体组分以及存在于靶上的甲基丙烯酰基。通过锚定在凝胶中的特定位点而固定在空间中的荧光标记所提供的位置信息现在可以在用蛋白酶进行化学预处理并随后在水中透析至延伸的构象之后物理地扩展。ExM的第一个描述是体积膨胀了100倍,靶分子之间的距离线性增加了4 - 5倍,使得SRM在传统共聚焦显微镜下的横向分辨率为70 nm,轴向分辨率为200 nm。在接下来的几年里,开发了改进的方案将蛋白质锚定到凝胶基质上而使得能够使用常规荧光团。为了避免自由基起始剂对荧光团的影响,蛋白质组放大分析等新方法(MAP)设计为与十二烷基硫酸钠配合使用(SDS)与热介导变性结合并能够对扩增的凝胶进行后标记。此外,已经引入了三官能接头,其在聚合,消化,并且能够将靶分子和官能团直接共价锚定到水凝胶中,从而能够使膜和细胞骨架元件可视化。最初的报告显示,一些荧光团会与自由基起始物发生反应。例如,花青不适合于当前的ExM方案,因为它们的信号在处理期间几乎消失。然而,其他荧光团(如Alexa Fluor 488、CF和Atto染料)在该方案中存活下来,并仍能发射足够的光子。此外,生物素-抗生物素蛋白信号放大和通过交换反应(Immuno-SABER)进行信号放大的免疫染色可有利地用于提高信噪比并对大量蛋白质靶点进行成像。最近研究表明,可以通过优化Ultra(U-ExM)保存中心粒的超微结构细节。U-ExM受到MAP方案的启发,MAP方案允许在扩增后进行后标记,并使用低甲醛和丙烯酰胺浓度小心交联蛋白质,以保留样品的超微结构细节。U-ExM已经成功地用于揭示以前只能通过电镜获得的中心粒的超微结构细节。通过使用不同的单体和引发剂组合,Truckenbrodt及其同事设计了一个方案,用于常规细胞培养物甚至神经元制备物的10倍扩增。如今,ExM已成功用于培养细胞、原代神经元和组织中的蛋白质、RNA、真菌、病理标本和细菌的超分辨率成像。4倍膨胀的ExM已经与晶格光片显微镜相结合,以60×60×90 nm的分辨率对整个果蝇大脑中多种蛋白质之间的纳米级结构进行成像(图4)。图4.ExM和点阵光片显微镜(LLSM)的结合可以显示大体积神经元组装的分子细节左上面板:概览图像,从右上角开始:海马CA1区锥体神经元投射;右下角:神经元体细胞器;底部中心:通过成像Thy1-EYFP信号结合突触蛋白的免疫组织化学可视化的小鼠脑、皮质树突棘;左下角:果蝇投射神经元(PN)结的多样性。左上:果蝇,PN至中枢复合体的神经元示踪。中心:果蝇,多巴胺能神经元的全脑可视化。底部面板:对成年果蝇脑的所有33个脑区域绘制的多巴胺能神经元相关Brp信号(体积密度)的定量(通过nc82抗体免疫染色,绿色曲线=与多巴胺能神经元相关的nc82点的百分比)。右上面板:初级体感皮质中沿皮质层I至VI的Homer 1点密度(顶部,最小强度投影)和皮质中25 μ m × 50 μ m × 50 μ m范围内Homer 1点的累积数量。注意II/III和V层中Homer 1的较高密度。为了进一步提高分辨率,ExM已经成功地与SRM方法相结合。例如,ExM与SIM结合,以20 nm的空间分辨率阐明果蝇和小鼠精母细胞中的联会复合体。ExM与STED在细胞培养样品中进行多表位标记后,与传统荧光显微镜相比,分辨率提高了30倍。此外,使用NHS染料结合物标记所有蛋白质的总和,然后进行迭代ExM,可以对高蛋白密度的结构或细胞器(如线粒体),实现与EM相比具有更高对比度的超微结构细节。为了在分子尺度上进行成像,ExM与SMLM方法(例如dSTORM)的组合是一个有吸引力的选择。通过使用低离子强度缓冲液成分或通过重新包埋到中性凝胶中稳定凝胶,可以避免收缩。另一种策略是使用自发闪烁荧光团(如HMSiR 137)在水中进行SMLM。通过Ex-dSTORM实现分子分辨率的关键是膨胀后标记,这增加了表位的可接近性并因此增加了标记效率并减少了连锁错误。Ex-dSTORM超分辨成像已成功应用于原代细胞和神经元中微管和中心粒结构的解析。4.1. 荧光标记 神经元和脑切片的超分辨率成像依赖于用合适的荧光团标记感兴趣的生物分子,理想地以定量和化学计量的方式。虽然SIM和所有其它超分辨率成像方法的性能取决于信号背景(S/B)比,但SIM对荧光团没有特殊要求。另一方面,STED显微镜可实现的分辨率主要取决于所用荧光团的光稳定性。RESOLFT显微镜使用具有两个稳定状态的可逆光开关FP,因此可使用较低的辐照强度。所有SMLM方法的定位精度取决于每个事件检测到的光子数。dSTORM需要光开关有机荧光团,包括花青、罗丹明和恶嗪染料,而PALM依赖于使用光开关、光转换和光活化FP。6与此相反,DNA-PAINT理论上适用于所有荧光团,因为开/关速率由对接和成像仪链序列和缓冲液条件决定,其中Cy3B和ATTO643迄今为止性能最佳。然而,独立于所使用的成像方法,具有最小连接误差(荧光团和靶之间的距离)的荧光探针的有效和特异性标记控制可获得的图像质量。为了通过荧光成像解析结构,标记密度(即荧光探针之间的距离)必须显著高于所需分辨率。另一方面,特别是对于接近实际分子分辨率的超分辨率成像方法,连接误差应尽可能小,以实现控制有效可实现分辨率的高成像精度。对于活细胞标记,在合适的表达载体中与目的蛋白融合的遗传编码FP无疑是选择的荧光团。然而,FP亮度较低,因此与有机染料相比,图像分辨率明显较低。理想的标记方法使用遗传编码的蛋白质、肽标签或可以用明亮荧光染料标记的单个氨基酸。由于瞬时转染产生不同蛋白表达水平的细胞,蛋白的分布和功能不一定反映野生型的情况。然而,构建体优化或稳定表达或CRISPR工程化敲入可产生所需的蛋白质水平,范围从内源性水平直至强烈过表达水平。根据策略,不同种类的转染方法可用于将核酸递送到神经元。传统的磷酸钙共沉淀法和脂质转染法遵循直观的验证方案,大多数实验室都有。不幸的是,这两种技术的转染效率较低。与此相反,病毒转导提高了效率,并允许注射到靶脑区域,但需要劳动密集型病毒生产方面的专业知识,并需要考虑增加生物安全水平法规。此外,应用病毒构建体时,必须考虑插入片段大小的相容性、毒性和差异表达起始。使用核转染实现最高的转染产率,其中施加高电压脉冲以将核酸直接递送到核。然而,当使用小鼠原代神经元时,细胞存活率较低,并且需要昂贵的设备,该设备还需要根据神经元密度和用于培养的动物种类对脉冲参数进行广泛的测试。通过在细胞附着结构中应用高电阻膜片吸管,在完整网络中例如在器官型切片中进行单细胞电穿孔也是可行的。基于这一知识,通过最近的基于crispr的基因敲入方法获得了接近内源性的蛋白质水平。在这里,在神经元发育的不同时间点,使用脂质转染、核转染或病毒递送在神经元中实现了基于卷曲的敲入。另一方面,FP表现出较低的光稳定性和荧光光子输出,从而降低了图像质量。此外,蛋白质功能可能会因附着大小为2 − 5 nm的FP而恶化。因此,必须检查目标蛋白质的野生型功能性。有机染料比FP小得多,并且表现出更高的光子产率和光稳定性,但是它们需要与结合到感兴趣的分子的其它分子偶联。对于固定细胞,使用一抗和二抗的免疫染色仍然是内源性蛋白标记的首选方法。然而,这导致了一个由两个IgG抗体的大小定义的连锁错误,即约17.5 nm。此外,在非常密集的细胞区室(如突触囊泡)中,用抗体对蛋白质进行有效的免疫标记仍然是一个非常具有挑战性的问题。通过使用初级荧光团标记的抗体或Fab片段可以减少连接错误。或者,针对GFP或基因导入短肽标签的更小(1.5 × 2.5 nm)骆驼抗体(纳米抗体)已用于dSTORM成像。8此外,荧光标记单体链霉亲和素可用于神经元和器官型组织中靶蛋白的特异性标记。使用这种标记方法,研究了神经肽-1 β、神经木质素-1和富含亮氨酸重复跨膜蛋白2的动力学和纳米级组织,并揭示了跨突触粘附结构的形成(图5)。图5.使用单体链霉亲和素结合AP-标记的突触蛋白通过dSTORM成像揭示神经木质素1和LRRTM2的不同分布上排:Homer1c-GFP作为突触后区室的参照。第二行:神经木质素1(Nlg1)和LRRTM2的dSTORM成像。底部,左侧:频率分布直方图,用于相对于导引头1信号的质心的信号色散的可视化。右下角:比较两种蛋白质的突触结构域数量的直方图。经参考文献159许可转载。版权所有2016施普林格自然。或者,可使用生物正交肽基序或自标记蛋白标签,例如FlAsH标签、SNAP标签和Halo标签。这些标签在基因上与靶蛋白共表达,并与各自的荧光团标记试剂或配体共价结合。对于肌动蛋白和微管丝的标记,可以使用小肽药物,如双环七肽鬼笔环肽和紫杉醇(紫杉烷类药物)。细胞膜和细胞器的标记可通过细胞器追踪器和荧光脂质类似物来实现。此外,小肽或配体可直接用荧光团标记,并特异性结合生物分子,例如,可显示抑制性突触后位点的超结合肽。然而,使用单个非天然氨基酸的位点特异性标记可实现最小的连接误差。在这里,通过遗传密码扩展引入定制设计的非天然氨基酸,并通过生物正交点击化学用四嗪染料标记。显然,神经元和组织切片必须根据要成像的结构进行透化和固定。无论使用何种标记方法,都必须特别注意所用化学品在其天然细胞环境中保存生物分子的超微结构。化学固定使蛋白质交联,可能对结合亲和力产生负面影响,也可能破坏分子间的相互作用。在大多数情况下,多聚甲醛(PFA)和戊二醛已成功用于神经科学的超分辨率成像。此外,还引入了乙二醛等新固定剂。膜分子应始终使用4% PFA和0.2%戊二醛固定,以最大限度地减少残留迁移率并避免人为现象,例如抗体结合诱导的簇形成。4.2. 神经元的多色遗传标记荧光蛋白质已经彻底改变了神经元的活细胞成像,因为它们可以通过基因融合到感兴趣的蛋白质上,并且在野生型功能的前提下,可以用于双色和三色成像。具有非常高密度的轴突和树突相互作用结构的神经系统需要用许多不同的颜色来标记以分离神经元连接。这个问题在2007年被称为Brainbow的转基因策略的发展所解决,该策略能够对神经元进行多色标记。结合单细胞分辨率成像,Brainbow可以用来创建大脑地图,这是一个连接体,详细描述了神经元如何形成回路以及它们投射到哪里。Brainbow策略利用了这样一个事实,即可见光谱中的所有颜色都可以由红、绿、蓝三原色的不同混合物产生。(RGB)或翻译成荧光蛋白,例如RFP、YFP和CFP。为了利用这一思想,广泛使用的Cre/lox重组系统(其可以通过DNA切除、倒位或染色体间重组来开启基因表达)已经被用于其第一次实现。这里,三种荧光蛋白基因中的一种在转基因中随机表达。多拷贝转基因盒的引入导致在每个细胞中具有不同拷贝数的三种基因的组合表达,从而产生几十种颜色,使得能够区分相邻神经元并可视化它们的相互作用。Brainbow技术非常适合绘制不同神经元类型之间的连接模式,追踪轴突,并识别大脑内长距离的神经元连接。此外,已证明Brainbow表达可成功用于研究周围神经损伤后的再生轴突,并检测脑发育过程中的重要步骤。为了进一步改善Brainbow在脑和包括突触蛋白在内的连通性绘图中的应用,SRM的应用是显而易见的,最近通过结合Brainbow、连续免疫染色、和ExM同时研究同一脑切片的形态学、分子标记物和连通性(图6)。将该技术转移到全脑研究一直是一个长期存在的障碍,直到最近才被解决。图6.结合Brainbow和ExM的多轮免疫染色和ExM(miriEx)成像(A)实验策略:在小白蛋白cre/+小鼠的脑切片中,在下一轮成像中,通过Brainbow信号和应用4 × ExM使小白蛋白阳性中间神经元可视化。突触通过Homer1和Gephyrin的免疫染色可视化,使用EYFP信号进行图像排列。(B)脑弓信号的免疫染色。(C)分别通过突触后标记homer1和Gephyrin的免疫染色区分抑制性和兴奋性突触。插图(D)−(F)和(G)−(I)显示了叠加图像的更高细节视图。(J)以及(K)神经元的形态重建(使用ImageJ插件nTracer),包括表征它们各自的输入。虚线框表示在(B)和(C)中描绘的区域。重建的神经元被连续编号。比例尺(扩展前):10微米((B)、(C))、2.5微米(I)、20微米((J)、(K))。经参考文献84许可转载。版权所有2020施普林格自然。4.3. 神经科学中的光电联合显微镜电子显微镜(EM)和电子断层扫描提供了细胞超微结构和细胞器组织的详细信息,其空间分辨率是光学显微镜无法达到的。然而,EM和电子断层摄影术不提供特异性分子标记,使得难以鉴定未知的细胞结构或表现出相似形态学特征的结构。用金结合抗体免疫标记EM能够以纳米分辨率定位蛋白质,但存在抗原标记效率低的问题,这意味着金颗粒的数量仅占抗原数量的1%至20%。另一方面,荧光显微镜提供大视场成像和活细胞中蛋白质的定位,但分辨率较低。在固定的样品中,用荧光团标记的抗体进行免疫标记使得能够对细胞中的各种分子进行有效和特异性的分子标记,并且与超分辨率荧光显微镜方法结合,空间分辨率远低于衍射屏障。因此,相关光电子显微镜(CLEM)是一种多功能的方法,可以在电子显微镜提供的细胞超微结构背景下通过超分辨率成像可视化蛋白质的定位和相互作用。然而,将超分辨率成像与EM相结合更困难,因为乍一看似乎主要是由于两种方法的不同和不相容的样品制备协议。例如,在EM中保存超微结构所需的固定和染色可引入强烈的自体荧光。此外,荧光蛋白被固定和聚合物包埋所需的脱水和氧化条件淬灭。此外,两个图像必须以纳米范围的精度重叠。这需要在荧光成像和EM中都表现出优异对比度的固定对准标记,例如裸金珠。此外,在荧光成像之后由样品脱水引起的结构变形会严重地损害两个图像的正确重叠。最终,必须在超微结构和荧光保存之间找到一个折衷方案。例如,对于某些周期性分子结构,如核孔复合体,dSTORM和扫描EM图像可以在不使用对准标记的情况下以〈20 nm的精度叠加。已经开发了能够在轴突和树突先前已经用荧光成像实时成像之后用透射EM观察轴突和树突的方案。为此,表达GFP的脑组织在荧光成像后被化学固定,并用电子致密标记物(例如用于EM的金)进行免疫标记。或者,荧光探针和附着于抗体的量子点的光氧化可用于补充更成熟的方法,例如过氧化物酶或胶体金标记。最后,二氨基联苯胺(DAB)聚合物可以通过光转化在荧光团处局部产生。为了克服标记问题并确保超微结构保存,已经开发了用于体积EM(NATIVE)的纳米体辅助组织免疫染色。NATIVE可有效标记蛋白质,无需苛刻的透化步骤、特殊树脂、锇替代品或清除剂。随着这些进步,CLEM已被证明非常适合绘制异质突触群和定位突触处的突触蛋白。参考文献(略)目前在国内,随机光学重建显微镜STORM已成功实现商用,有需要STORM成像技术进行实验研究的专家老师们,可预约获得 iSTORM 超高分辨率显微成像系统试拍服务~图、iSTORM拍摄的超高分辨率细胞图片力显智能现已发布的超高分辨率显微成像系统 iSTORM,成功实现了光学显微镜对衍射极限的突破,使得在20纳米的分辨率尺度上从事生物大分子的单分子定位与计数、亚细胞及大分子复合物结构解析、生物大分子生物动力学等的研究成为现实,从而给生命科学、医学等领域带来重大突破。关于我们About us 宁波力显智能科技有限公司(INVIEW)是专业从事超高分辨率显微技术和产品研发的科技企业,依托复旦大学的自动控制、新一代信息技术及香港科技大学的生物、光学、图像处理等的技术,拥有光学、生物、自控、机械、信息技术等多领域交叉学科技术团队,将2014年诺贝尔化学奖技术产业化,推出了超高分辨率显微成像系统iSTORM、细胞智能监控助手赛乐微等一系列产品,帮助人们以前所未有的视角观察微观世界,突破极限,见所未见。
确定

产品配置单

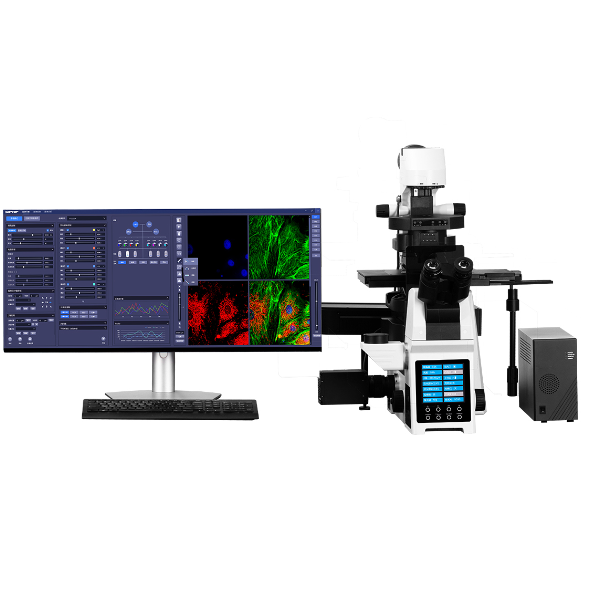
宁波力显智能科技有限公司为您提供《【预约试拍】超高分辨显微镜在神经科学中的应用(二)》,该方案主要用于其他中超高分辨率显微镜检测,参考标准--,《【预约试拍】超高分辨显微镜在神经科学中的应用(二)》用到的仪器有超高分辨显微镜iSTORM
推荐专场




相关方案
更多