








方案详情
文
WIPM NMR 波谱仪检测数据的分析结果与 Bruker NMR 波谱仪的实验结果具有高度的一致性,表明 WIPM 谱仪可以用于代谢组学研究.研究发现减毒鼠伤寒沙门氏菌感染能够引起小鼠粪样代谢物中多种氨基酸和尿嘧啶含量的上升,并伴有胆汁酸、乳酸和丙酸含量的下降.以上代谢物含量的改变表明,减毒鼠伤寒沙门氏菌感染使小鼠肠道微生物的代谢功能发生了紊乱.
方案详情

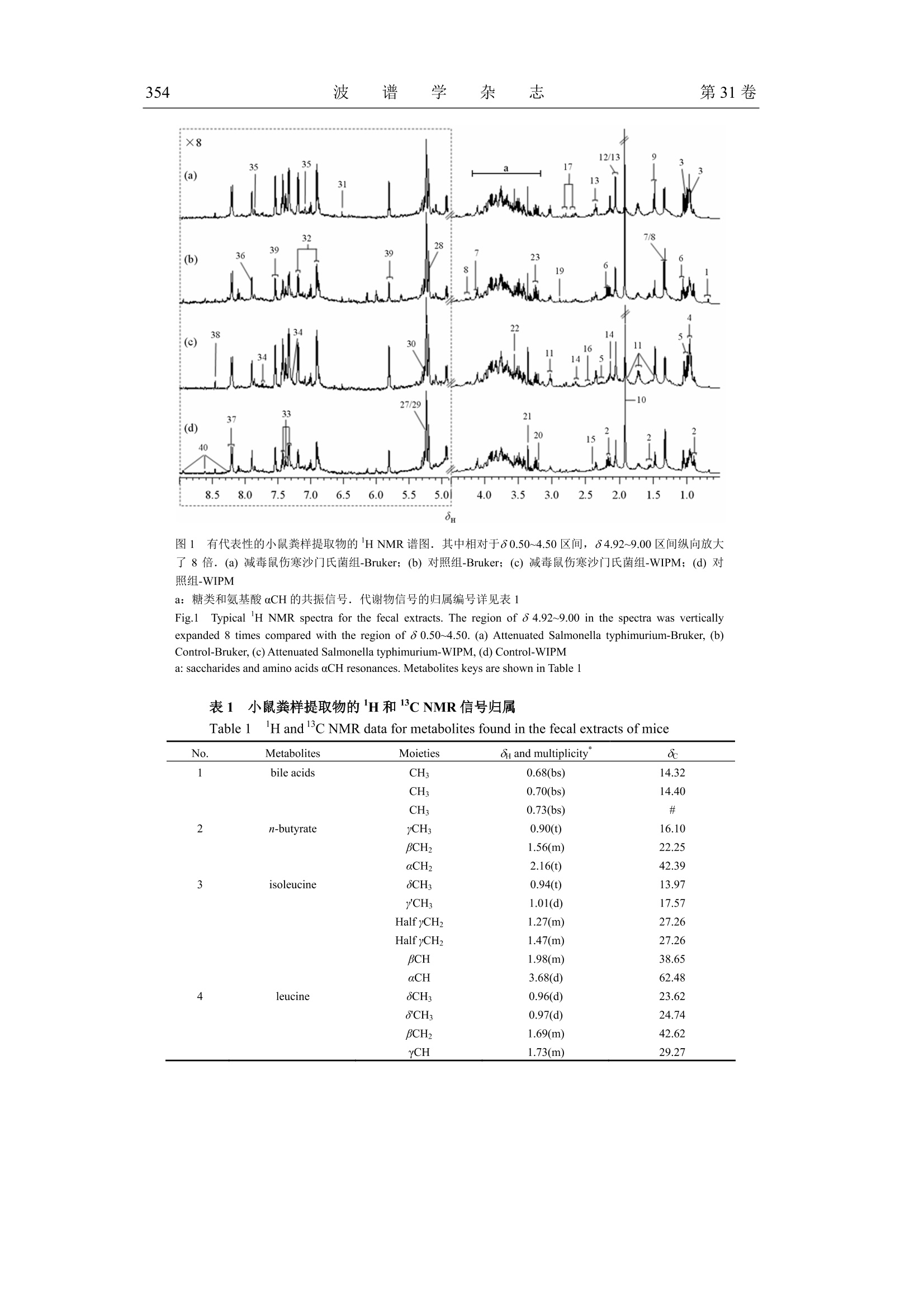
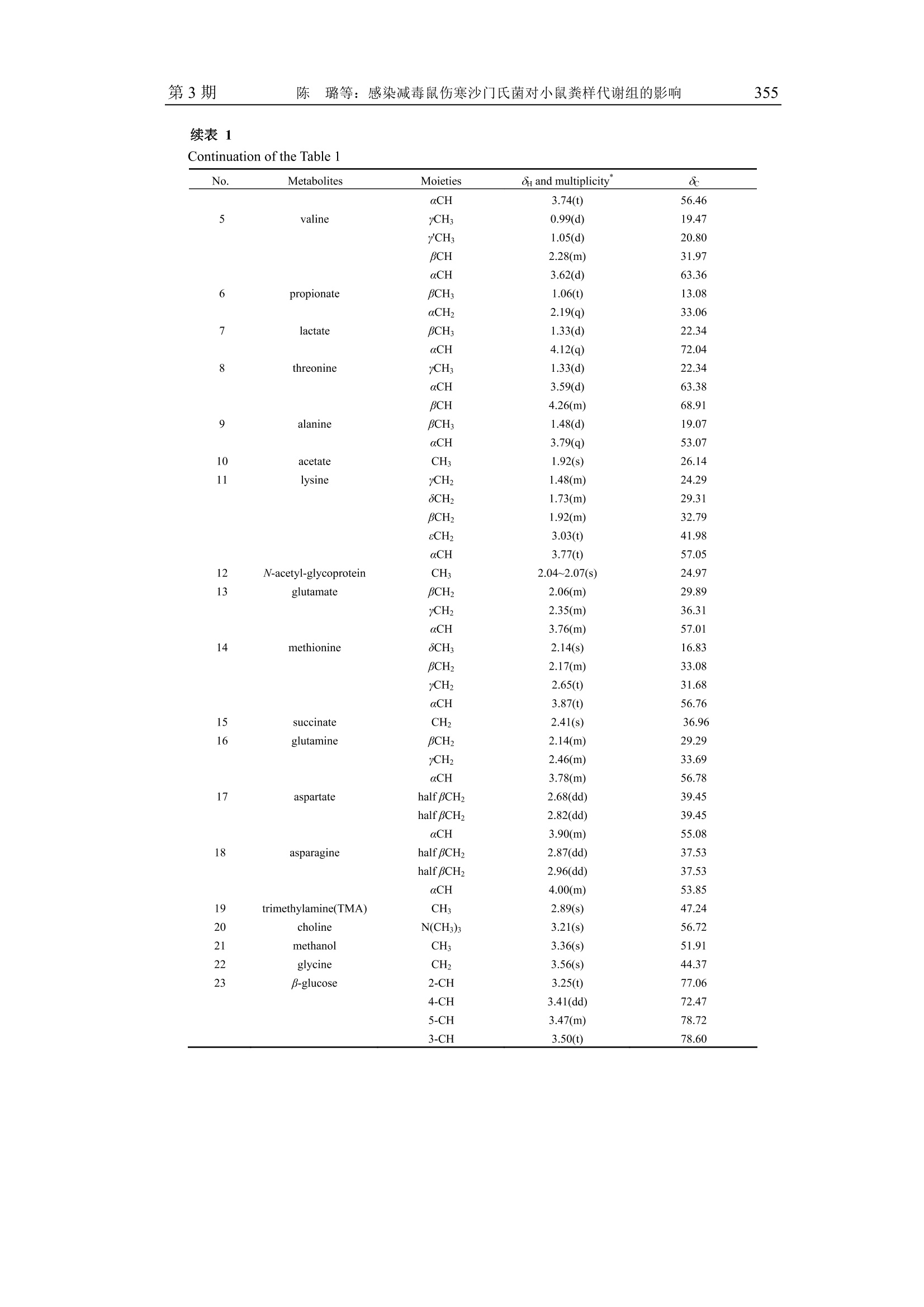
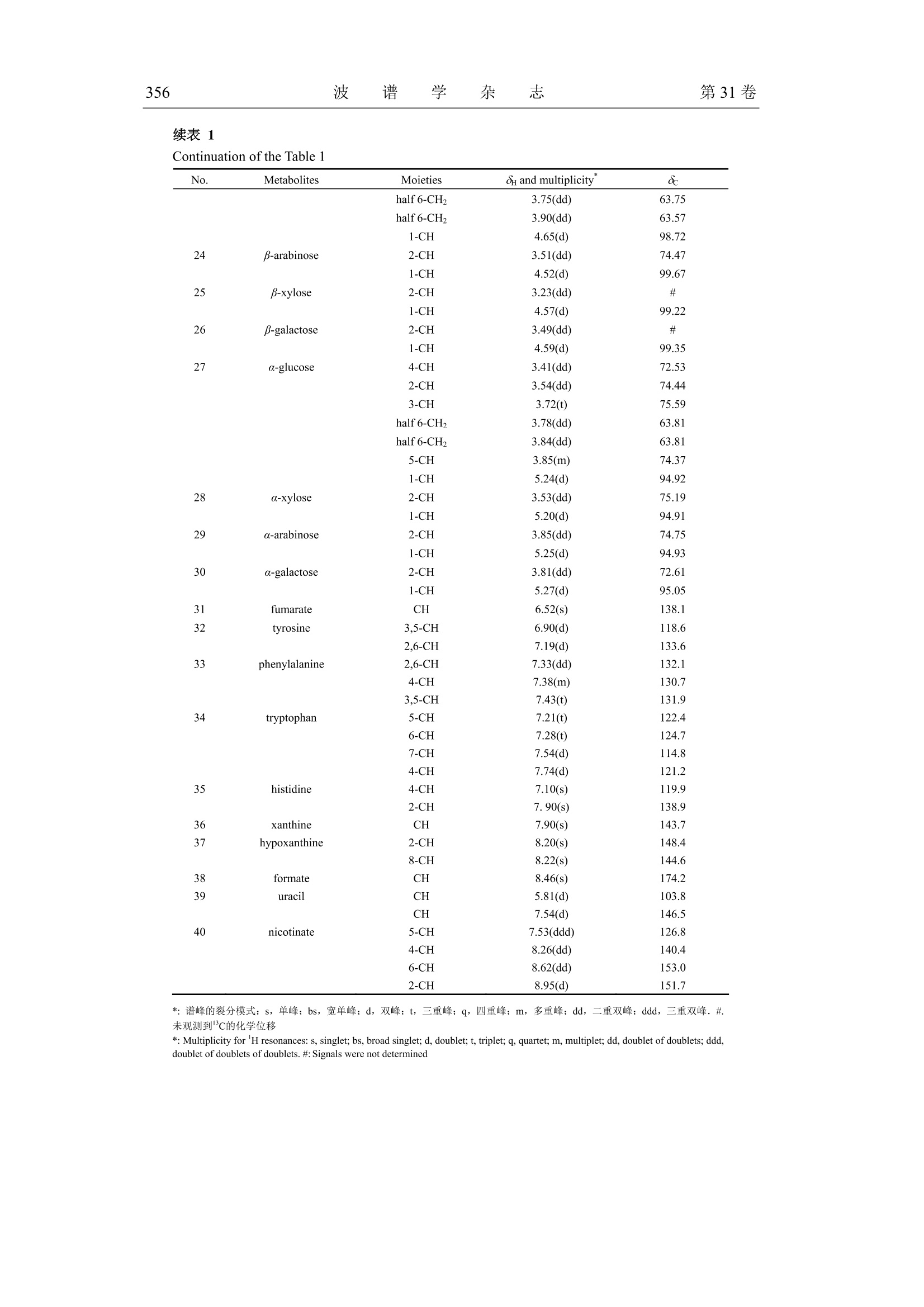
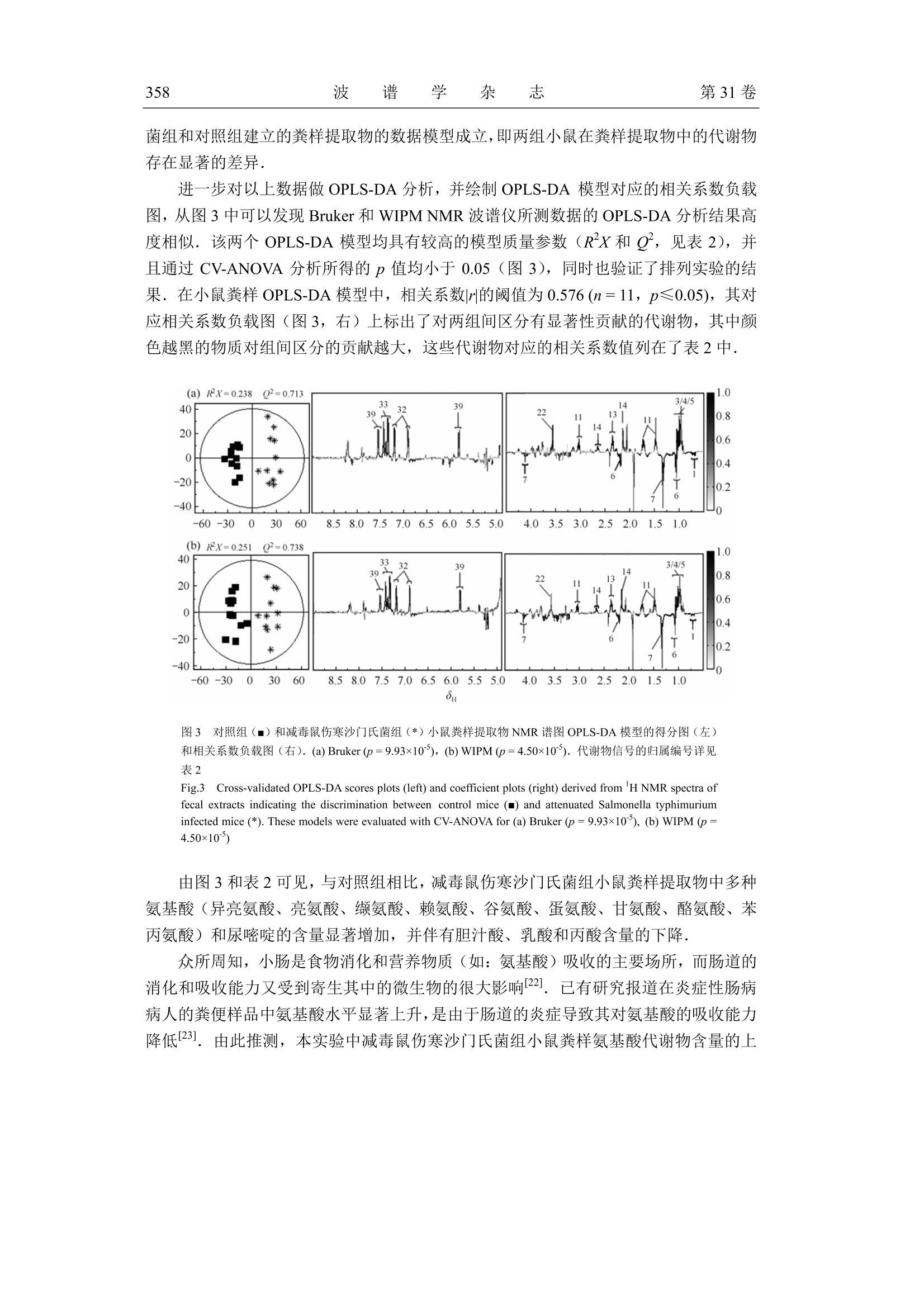
波 谱 学 杂 志Chinese Journal of Magnetic ResonanceVol. 31 No. 3Sep. 2014第31卷第3期2014年9月 波 谱 学 杂 志第31卷350 文章编号:1000-4556(2014)03-349-15 感染减毒鼠伤寒沙门氏菌对小鼠粪样代谢组的影响 -WIPM 和 Bruker 500 MHz 核磁共振波谱仪检测结果的比较 陈 璐1.2宋 侃品113,,王玉兰1* [1.中国科学院生物磁共振分析重点实验室,波谱与原子分子物理国家重点实验室,武汉磁共振中心 (中国科学院武汉物理与数学研究所),湖北武汉430071;2.长江航务管理局疾病预防控制中心,湖北武汉430019; 3.中国科学院大学,北京100049] 摘 要:鼠伤寒沙门氏菌属于胞内侵袭性兼性厌氧菌,减毒后的鼠伤寒沙门氏菌安全性大大提高,已被广泛用于病毒、细菌及寄生虫疫苗的研制.但是目前还没有研究利用代谢组学方法来分析减毒鼠伤寒沙门氏菌感染对宿主和肠道菌群相互作用的影响.同时中国科学院武汉物理与数学研究所自主研发的 WIPM 500 MHz 核磁共振(NMR)波谱仪已经研制成功,但是该谱仪是否可以用于代谢组学研究尚未经过考证.本研究同时使用 WIPM 和 Bruker500 MHz 核磁共振波谱仪对同一批减毒鼠伤寒沙门氏菌感染小鼠的粪样进行检测,并结合多变量数据分析方法来研究减毒鼠伤寒沙门氏菌感染对小鼠粪样代谢组的影响.研究结果表明, WIPM NMR 波谱仪检测数据的分析结果与 Bruker NMR 波谱仪的实验结果具有高度的一致性,表明 WIPM 谱仪可以用于代谢组学研究.研究发现减毒鼠伤寒沙门氏菌感染能够引起小鼠粪样代谢物中多种氨基酸和尿嘧啶含量的上升,并伴有胆汁酸、乳酸和丙酸含量的下降.以上代谢物含量的改变表明,减毒鼠伤寒沙门氏菌感染使小鼠肠道微生物的代谢功能发生了紊乱. 关键词:核磁共振(NMR);粪样代谢组;减毒鼠伤寒沙门氏菌;谱仪对比 中图分类号:O482.53 文献标识码:A ( 收稿日期:2014-01-14;收修改稿日期:2014-07-21 ) ( 基金项目:国 家 重大科学仪器设备开发专项资助项目(2011YQ12003503). ) ( 作者简介:陈璐(1988-), 女 ,江西抚州人,硕士研 究 生,主要从事生物医学代谢组学研究, E-mail: tjchenlu@163.com. *通讯联系人:王玉兰, 电 话:027-87198 4 30, E-mail: yulan. w ang@wipm.ac.cn. ) 引言 鼠伤寒沙门氏菌(Salmonella typhimurium)是目前分离率极高的一种沙门氏菌(Salmonella)菌型,同时也是重要的人兽共患病原菌,其不仅能够引起人与畜禽的自限性肠炎,还可以导致小鼠出现人伤寒样的系统性感染以及人类的食物中毒等多种症状!.有效疫苗的研制在沙门氏菌病的防控过程中扮演着极为重要的角色,越来越多的研究证明,减毒沙门氏菌活疫苗具有比亚单位疫苗或死疫苗更安全、更有效的免疫效果[2.所谓减毒沙门氏菌是指通过物理、化学或基因工程等手段,使沙门氏菌某些特定的基因发生不可逆的突变,进而使其毒性大大降低.减毒后的鼠伤寒沙门氏氏安全性性大提高,并在降低致病力的同时仍保持良好的侵袭力和免疫原性.减毒鼠伤寒沙门氏菌通过口服途径感染宿主后,能够产生持续且有效的细胞、体液以及局部黏膜免疫效应,目前已作为疫苗及 DNA 疫苗载体被广泛用于细菌[3,4]、病毒I5,6]及寄生虫[7-9]疫苗的研制. 近几十年来,随着鼠伤寒沙门氏菌基因组学、蛋白质组学和代谢组学等组学研究的开展,人类开始从分子水平对鼠伤寒沙门氏菌的多种特性进行研究.其中代谢组学出现的时间相对较晚,但它在短短十几年里的发展及其在系统生物学研究中的重要性却不可小觑.代谢组学能够揭示生物体最终确实已经发生了的变化,并协助研究者以下游代谢产物为出发点,进一步研究生命过程的本质及特性[10],其常用的检测方法有: NMR、色谱(高效液相色谱法, HPLC;气相色谱, GC)、质谱(MS)以及多种技术的联用1.目前,多数研究都是以沙门氏菌培养基为对象进行的代谢组学研究,通过研究结果间接反映出菌体自身所发生的代谢变化[12].后来有研究者通过傅立叶变换离子回旋共振(DI-FT-ICR) MS来分析鼠伤寒沙门氏菌对小鼠肝脏和粪样代谢轮廓的影响,研究结果显示鼠伤寒沙门氏菌扰乱了宿主体内的多条代谢途径,其中激素类物质(类花生酸、类固醇等)发生的变化最为明显[13].然而,在这些工作中与 NMR 相关的报道极少, NMR检测技术仅被用于鼠伤寒沙门氏菌及其减毒菌株中某些特定物质(蛋白、酶类及脂质 A等)的结构分析,迄今还没有研究利用 NMR 技术来分析减毒菌株感染对宿主代谢组产生的影响. 本研究的对象为减毒鼠伤寒沙门氏菌 YS14510 (purl, msbB),其绝大部分特性与VNP20009 (purl, msbB , xylose )这种常见减毒菌株相似,VNP20009 的相关研究表明,该种减毒菌株具有极佳的安全性,并且在体内、外经过多次传代(>140)仍能保持遗传性状的稳定性.此外, VNP20009 在小鼠体内的清除率非常快,在感染后24h便无法在尿液和粪便中检测到菌体的存在;在组织中则优先聚集在肝脏和脾脏当中,菌体数目随着时间的推移而明显减少,最终于3个月后被全部清除[14,15]然而这些研究仅仅关注了减毒菌的使用安全性,却忽略了减毒鼠伤寒沙门氏菌在排出宿主体外以后,尤其是从粪便中排出后,其对宿主代谢应答产生的潜在效应. 基于 NMR 的代谢组研究能够充分利用NMR 技术的无选择、无偏向、无损伤、灵活以及高通量等特性,对生物代谢物质进行定性定量分析[16].在全球的分析仪器生产 商中,德国 Bruker-Biospin 公司一直占据着该领域的领先地位.为了将中国科学院武汉物理与数学研究所(WIPM)自主研发的 NMR波谱仪与 Bruker NMR 波谱仪的检测性能进行比较,同时开展基于自主研发 NMR 波谱仪的代谢组学研究,本文同时使用 WIPM 和Bruker 500 MHz NMR 波谱仪对同一批减毒鼠伤寒沙门氏菌感染小鼠的粪样进行检测,并结合多变量数据分析方法来研究减毒鼠伤寒沙门氏菌对小鼠粪样代谢组的影响,旨在为减毒鼠伤寒沙门氏菌的研究工作提供基础的粪样代谢组学信息,同时为 WIPM NMR波谱仪的产业化提供可靠的科研依据. 实验部分 1.1 菌株及实验动物 减毒鼠伤寒沙门氏菌 YS14510 由美国亚利桑那州立大学生命科学院施一燊教授惠赠.订该菌株通过基因工程手段,敲除了野生型鼠伤寒沙门菌(ATCC 14028)的 msbB 和purI 基因而获得.24只无特定病原体(Specific Pathogen Free,SPF)级 6周龄雌性 BALB/c小鼠(20±2g),购于湖南斯莱克景达实验动物有限公司,实验动物合格证编号:HNASLKJ20120137. 1.2 化学试剂 重水(D20,99.9%氛代)购于美国 Sigma-Aldrich 贸易有限公司;胰蛋白胨、酵母提取物和琼脂粉购于英国 OXOID 公司,用于配制 Luria-Bertani(LB)培养基;2,2,3,3-氛代三甲基硅烷丙酸钠(TSP,98%氛代)购于美国 Cambridge Isotope Laboratories 公司;分析纯叠氮钠(NaN3)购于天津福晨化学试剂厂;分析纯氯化钠(NaCl)、氯化钾(KCl)、磷酸氢二钾(K2HPO4·3H2O)和磷酸二氢钠(NaH2PO4·2H2O)购于上海国药集团试剂有限公司. 1.3 实验方法 1.3.1 磷酸盐缓冲溶液(Phosphate Buffered Saline, PBS)的配制 缓冲液A(K2HPO4/NaH2PO4, 0.01 mol/L, pH=7.4)由双蒸水(ddH2O)配制,其中含有0.8%NaCl (w/v)和 0.02%KCl (w/v),经121℃高温高压灭菌 20 min 后4℃保存,用于菌悬液的制备.粪样提取使用的缓冲液 B(K2HPO4/NaHPO4, 0.1 mol/L, pH=7.4)由 50%D2O配制,其中含有 0.002% TSP (w/v)和 0.1% NaN3 (w/v)分别作为化学位移内标物和防腐剂. 1.3.2 细菌的培养及菌悬液的制备 取出-80℃冻存的 YS14510菌株,在室温下融解后,接种1-2环菌液于LB平板上连续划线培养.37℃活化培养24h 后,挑取单菌落转接于 5 mL LB 液体培养基中,37℃振荡培养过夜,次日分别取200 uL 培养液接种于两块 LB平板,涂布均匀后在37℃下培养.24h后刮取菌落(不可刮入培养基),混悬于 PBS中制备成 YS14510 菌悬液,并将菌悬液浓度调至1×10°菌落形成单位(Colony-Forming Units, CFU)/mL. 1.3.3 动物实验方案及样品的收集 适应性饲养2周后,将24只 SPF 级 BALB/c 小鼠随机分为减毒鼠伤寒沙门氏菌组和对照组,每组各12只.减毒鼠伤寒沙门氏菌组每只小鼠灌服 YS14510 菌悬液0.3 mL(1x10°CFU/mL),每隔2周一次,连续3次.对照组以0.3mL 无菌PBS代替,每隔2周灌服一次,连续3次.所有动物行灌胃处理前禁食禁水4h,末次灌喂2周后收集所有小鼠粪样.粪样一经收集立即以液氮速冻,然后置于-80℃保存待 NMR 检测. 1.3.4 粪样的提取及制备 称取 50 mg 左右小鼠粪样置于2mL离心管中,加入 0.5 mL 粪样提取缓冲液 B,液氮反复冻融3次(于25℃水浴中解冻,解冻完成后涡旋混匀),再用组织研磨仪(Tissuelyser, QIAGEN, 德国)进行匀浆(20Hz, 90 s), 然后对匀浆液进行离心(10000g,4℃,10 min)取上清.对剩余沉淀用相同方法再重提2次,最后将3次上清液混合后再离心(16000 g, 4℃, 10 min), 取上清 550 uL 转移至 5 mm NMR 样品管中待 NMR 检测1. 1.3.5 样品的NMR检测 本实验同时应用 WIPM-I 500 型NMR波谱仪(中国科学院武汉物理与数学研究所自主研发,质子共振频率为 499.77 MHz) 和 Bruker AVII 500 型 NMR 波谱仪(德国Bruker-Biospin 公司,质子共振频率为 500.13 MHz) 对同一批小鼠粪样采集 'HNMR谱图,实验温度为 298 K. 1D H NMR 实验所用的脉冲序列为 Noesygpprld [RD-G-90°-ty-90°-tm-G2-90°-ACQ],具体的采样参数设置如下:循环等待时间(RD)为2s, 混合时间(tm)为 100 ms,脉冲延迟时间(ti)为4 us,谱宽为10000Hz, 采样时间为1.64 s, 采样点数为32k,累加次数为64. 为了进一步对1D 谱中的信号进行归属,还需要选取具有代表性的样品采集一系列的 2D NMR 谱, 即'H-'HCOSY, H-'HTOCSY, H-HJ-RES, H-3C HSQC和'H-IcHMBC 五种谱图.所有2DNMR 实验在 Bruker AVIII 600 型 NMR 波谱仪上进行,该谱仪配有TXI超低温探头,其质子共振频率为 600.13 MHz.'H-'H J-RES 的F('H)维的谱宽为6313.13 Hz,采样数据点阵 t2xt=4096×64;在'H-H COSY与'H-'HTOCSY实验中F2(H)和F(H)维的谱宽均为6313.13Hz, 采样数据点阵 tzxtj=2048×128;'H-CHSQC的F('H)和F(C)维的谱宽分别为 6 313.13 Hz 和26409.97 Hz,采样数据点阵 tzxt=2048×90;H-13CHMBC的F(H)和F(C)维的谱宽分别为 6 313.13 Hz和33201.94 Hz, 采样数据点阵 txt=2048×100. 1.3.6 NMR 谱图处理及多变量数据分析 WIPM 500 型 NMR 波谱仪上采集到的 1D 'HNMR谱图通过 SpinStudio 软件(中国科学院物理与数学研究所自主研发)进行处理,Bruker 500 型NMR 波谱仪上采得的1D谱图则通过 Topspin3.0 软件(德国 Bruker-Biospin 公司研发)进行处理:所有自由感应衰减信号(FID)均乘以增宽因子为1Hz的指数窗函数,经傅立叶变换(FT)后对谱图进行手动相位和基线的校正,然后用内标 TSP 的单峰定标(80.00). 再使用 AMIX 软件 (V3.9.5,德国 Bruker-Biospin 公司)对 'H NMR 谱在80.50~9.00的区域进行分段积分,积分区间宽为0.004.为了消除残余水峰信号的影响,去除 NMR 谱中的84.50~4.92(水信号)区间. 对积分后的数据进行总面积归一化处理,然后导入 SIMCA-P+软件包(12.0,瑞典Umetrics 公司)进行多变量分析.首先主成分分析(PCA)采用中心化处理(Ctr)的标准化处理方式,得到样品整体的分布趋势,同时可以观察到组内有无异常点,分析结果用得分图(Scores plots)来表示18].然后使用偏最小二乘法判别分析(PLS-DA)发现 NMR 数据(X变量)和分组变量(Y变量)之间的相关关系,用舍一法对模型的质量进行交叉验证(Cross-validation, CV), 得到R’X(代表模型可解释的变量)和g(代表模型的可预测度),然后通过排列实验(Permutation test)和 CV-ANOVA19验证模型的有效度.最后以正交偏最小二乘法判别分析(OPLS-DA)最大化地凸显组别之间的差异,并利用MATLAB 7.1软件(美国Mathworks 公司)结合研究组自编程序作出相关系数负载图(Coefficient Plot), 结果显示为灰度图,即用黑白渐进色来表征代谢物对组间分分的贡献大小.根据两组中较小的样本数,查阅皮尔森相关系数(Pearson Correlation Coefficient)临界值表,得到相应的阈值 reutoff.当某一代谢物信号的相关系数绝对值|r≥rcutoff时,则认为该代谢物的变化对模型的区分有意义,颜色越黑,代表|r|越大,其对组间区分上的贡献也越大[20]. 2 结果与讨论 2.1 小鼠粪样提取物的HNMR谱图分析 图1是在 Bruker 和 WIPM 500 MHz NMR 波谱仪上采集的具有代表性的减毒鼠伤寒沙门氏菌组和对照组小鼠粪样提取物的 ‘H noesygppr1d NMR 谱图,从1DNMR谱图中可以获取样品中不同物质的化学位移、偶合常数、裂分情况以及谱峰面积比例等相关信息,其中根据化学位移可以初步判断质子所处的化学基团种类.同时,依据一系列2D NMR实验和相关文献211,并结合各种公共数据库对样品中的代谢物质进行谱峰的归属和物质的确认,详细的归属信息见表1. 从图1中观察到 Bruker 和 WIPM 500 MHz NMR 波谱仪上采集的 1D 'HNMR 谱图相似度极高,二者检测出的代谢物及峰强度大小均非常吻合.从谱图中得到小鼠粪便提取物中主要由氨基酸(异亮氨酸、亮氨酸、缬氨酸、苏氨酸、丙氨酸、赖氨酸、谷氨酸、蛋氨酸、谷氨酰胺、天冬氨酸、天冬酰胺、甘氨酸、酪氨酸、苯丙氨酸、色氨酸、组氨酸等)、短链脂肪酸(甲酸、乙酸、丙酸、正丁酸等)、糖类(葡萄糖、阿拉伯糖、木糖、半乳糖等)、核苷酸(如:黄嘌呤、次黄呤、尿嘧啶等)以及胆汁酸等代谢物组成.由肉眼观察上述谱图可见减毒鼠伤寒沙门氏菌组和对照组小鼠粪样提取物的代谢物组成存在一定的差异,如:减毒鼠伤寒沙门氏菌组小鼠粪便中异亮氨酸、亮氨酸、缬氨酸以及苯丙氨酸等氨基酸类物质含量相对较多,而对照组中含有较高水平的乳酸. 图1 有代表性的小鼠粪样提取物的HNMR谱图.其中相对于80.50~4.50区间,84.92~9.00区间纵向放大了 8倍. (a)减毒鼠伤寒沙门氏菌组-Bruker;(b) 对照组-Bruker; (c) 减毒鼠伤寒沙门氏菌组-WIPM;(d)对照组-WIPM a:糖类和氨基酸 αCH 的共振信号.代谢物信号的归属编号详见表1 Fig.1 Typical H NMR spectra for the fecal extracts. The region of 8 4.92~9.00 in the spectra was verticallyexpanded 8 times compared with the region of 8 0.50~4.50. (a) Attenuated Salmonella typhimurium-Bruker, (b)Control-Bruker, (c)Attenuated Salmonella typhimurium-WIPM,(d) Control-WIPM a: saccharides and amino acids aCH resonances. Metabolites keys are shown in Table 1 表1 小鼠粪样提取物的H和13CNMR信号归属 Table l 'Handc NMR data for metabolites found in the fecal extracts of mice No. Metabolites Moieties 8 and multiplicity 8c 1 bile acids CH: 0.68(bs) 14.32 CH 0.70(bs) 14.40 CH3 0.73(bs) # 2 n-butyrate yCH3 0.90(t) 16.10 BCH, 1.56(m) 22.25 aCH, 2.16(t) 42.39 3 isoleucine 8CH: 0.94(t) 13.97 y'CH: 1.01(d) 17.57 Half yCH2 1.27(m) 27.26 Half yCH2 1.47(m) 27.26 BCH 1.98(m) 38.65 aCH 3.68(d) 62.48 4 leucine SCH 0.96(d) 23.62 8'CH 0.97(d) 24.74 BCH2 1.69(m) 42.62 yCH 1.73(m) 29.27 续表1Continuation of the Table 1 续表 1Continuation of the Table 1 *:谱峰的裂分模式: s,单峰; bs, 宽单峰; d, 双峰; t,三重峰;q,四重峰; m,多重峰; dd,二重双峰; ddd, 三重双峰. #.未观测到C的化学位移 *: Multiplicity for H resonances: s, singlet; bs, broad singlet; d, doublet; t, triplet; q, quartet; m, multiplet; dd, doublet of doublets; ddd,doublet of doublets of doublets. #: Signals were not determined 然而肉眼观察谱图获取的信息十分有限,为了获取更多关于减毒鼠伤寒沙门氏菌引发小鼠粪样代谢物质变化的信息,本文进一步采用了多变刃数据分析方法对 NMR 数据进行深入的分析. 2.2 粪样提取物'HNMR 谱图的多变量分析 为了观察动物模型中样本的聚类情况以及是否存在离群点,本研究先分别对在Bruker 和 WIPM 500 MHz NMR 波谱仪上采集的小鼠粪样提取物的 NMR 数据进行 PCA分析.二者的 PCA 得分图[图2(a)和图 2(d)]中均显示对照组和减毒鼠伤寒沙门氏菌组小鼠粪样提取物的代谢物组成存在较明显的分离趋势.其中发现减毒鼠伤寒沙门氏菌组粪样提取物中有一个样品(编号:b9)为明显的离群点,通过观察原始谱图发现其所有代谢物的含量均明显高于同组其它样品,考虑为严重的个体差异所致,为了更准确地对数据进行解释,后续的数据分析过程中将该样品去除. 图2对照组()和减毒鼠伤寒沙门氏菌组(*)小鼠粪样提取物 'HNMR 谱图的 PCA 得分图、PLS-DA得分图及 PLS-DA模型的排列实验结果. (a) PCA 得分图-Bruker; (b) PLS-DA得分图-Bruker; (c)排列实验-Bruker; (d) PCA 得分图-WIPM; (e) PLS-DA 得分图-WIPM; (f) 排列实验-WIPM Fig.2 PCA scores plots, PLS-DA scores plots and permutation test for control mice () and attenuatedSalmonella typhimurium infected mice (*) derived from the H NMR spectra of fecal extracts. (a) PCA scoresplots-Bruker, (b) PLS-DA scores plots-Bruker, (c) permutation test-Bruker, (d) PCA scores plots-WIPM, (e)PLS-DA scores plots-WIPM, (f) permutation test-WIPM 在PCA 的基础上,接着进行 PLS-DA 分析, Bruker 和 WIPM NMR 数据的 PLS-DA模型验证的排列实验结果如图2(c)和图2(f)所示,实验结果均表明,减毒鼠伤寒沙门氏 菌组和对照组建立的粪样提取物的数据模型成立,即两组小鼠在粪样提取物中的代谢物存在显著的差异. 进一步对以上数据做 OPLS-DA分析,并绘制 OPLS-DA 模型对应的相关系数负载图,从图3中可以发现 Bruker 和 WIPM NMR 波谱仪所测数据的 OPLS-DA 分析结果高度相似.该两个 OPLS-DA 模型均具有较高的模型质量参数(RX和g,见表2),并且通过 CV-ANOVA 分析所得的p值均小于0.05(图3),同时也验证了排列实验的结果.在小鼠粪样 OPLS-DA模型中,相关系数|r的阈值为 0.576 (n=11,p≤0.05), 其对应相关系数负载图(图3,右)上标出了对两组间区分有显著性贡献的代谢物,其中颜色越黑的物质对组间区分的贡献越大,这些代谢物对应的相关系数值列在了表2中. 图3 对照组(■)和减毒鼠伤寒沙门氏菌组(*)小鼠粪样提取物 NMR 谱图 OPLS-DA 模型的得分图(左)和相关系数负载图(右). (a) Bruker (p=9.93×10*), (b) WIPM(p=4.50x10*). 代谢物信号的归属编号详见 表2 Fig.330Cross-validated OPLS-DA scores plots (left) and coefficient plots (right) derived from H NMR spectra offecal extracts indicating the discrimination between control mice (■) and attenuated Salmonella typhimuriuminfected mice (*). These models were evaluated with CV-ANOVA for (a) Bruker (p=9.93×10*), (b) WIPM (p=4.50×105) 由图3和表2可见,与对照组相比,减毒鼠伤寒沙门氏菌组小鼠粪样提取物中多种氨基酸(异亮氨酸、亮氨酸、缬氨酸、赖氨酸、谷氨酸、蛋氨酸、甘氨酸、酪氨酸、苯丙氨酸)和尿嘧啶的含量显著增加,并伴有胆汁酸、乳酸和丙酸含量的下降. 众所周知,小肠是食物消化和营养物质(如:氨基酸)吸收的主要场所,而肠道的消化和吸收能力又受到寄生其中的微生物的很大影响[22].已有研究报道在炎症性肠病病人的粪便样品中氨基酸水平显著上升,是由于肠道的炎症导致其对氨基酸的吸收能力降低[23].由此推测,本实验中减毒鼠伤寒沙门氏菌组小鼠粪样氨基酸代谢物含量的上 升,可能是因为小鼠感染减毒伤寒沙门氏菌后,引起了体内的炎症反应使其肠道微生物发生紊乱,最终导致小肠的吸收不良,并伴随粪样中氨基酸含量的升高.此外,本研究还观察到粪样中尿嘧啶含量增加,提示感染减毒鼠伤寒沙门氏菌所造成的早期炎症反应可能会导致小鼠组织细胞对 DNA 和 RNA 生物合成所需的嘧啶核苷酸需求增加,并使得与尿嘧啶降解速度密切相关的二氢嘧啶脱氢酶(DPD)活性下调[24],最终导致尿嘧啶分解速率减慢. 表2 对照组和减毒鼠伤寒沙门氏菌组小鼠粪样中发生显著变化的代谢物及其 OPLS-DA相关系数 Table 2 Correlation Coefficients for Metabolites with Significant Differences between controland attenuated Salmonella typhimurium infected mice Bruker 500 MHz WIPM 500 MHz Metabolites (No.) RX=0.238Q=0.713 R2x=0.251Q=0.738 r|≥0.576 r≥0.576 bile acids (1) -0.808 -0.767 Isoleucine (3) +0.804 +0.775 leucine (4) +0.788 +0.781 valine (5) +0.818 +0.803 propionate (6) -0.715 -0.672 lactate (7) -0.755 -0.808 lysine (11) +0.774 +0.749 glutamate (13) +0.719 +0.753 methionine (14) +0.767 +0.819 glycine (22) +0.787 +0.708 tyrosine (32) +0.791 +0.847 phenylalanine (33) +0.845 +0.889 uracil (39) +0.726 +0.749 t:+和-分别表示与对照组相比减毒鼠伤寒沙门氏菌组代谢物相对含量的增加和减少 t: The +and-signs indicate metabolite concentration increase and decrease respectively for attenuated Salmonella typhimurium infected mice compared with control mice 在正常生理状态下,90%的胆汁酸在肠道中通过肝肠循环被重吸收,而肠道菌群在胆汁酸的代谢过程中扮演着重要的角色,因此胆汁酸含量的变化同样也可以反应宿主-肠道微生物的相互作用[25].实验结果发现减毒鼠伤寒沙门氏菌组动物的粪样中胆汁酸含量显著下降,这表明减毒鼠伤寒沙门氏菌感染可能使肠道微生物代谢功能发生了紊乱,从而影响了胆汁酸的正常代谢. 短链脂肪酸(SCFAs)主要是由食物中半纤维素和抗性淀粉(Resistant starch)经肠道厌氧菌发酵生成的产物.它们不仅是动物体内的主要能量来源之一,还可以维护肠道上皮 细胞的完整性,对粘膜免疫细胞有维护作用,并保护肠道不受炎症的侵害[26].哺乳动物由于缺乏相应的酶类,所以必须依赖肠道菌群来参与代谢食物中这类不消化的碳水化合物[271.前人还发现肠道菌群可将乳酸转化为短链脂肪酸,这也是盲肠内容物中短链脂肪酸的来源之一[28].本研究中观察到减毒鼠伤寒沙门氏菌组小鼠粪样中乳酸和丙酸含量显著降低,代表在减毒鼠伤寒沙门氏菌感染后,小鼠肠道微生物对糖类物质的发酵功能受到了影响,进而使相应发酵产物—短链脂肪酸的含量降低. 由此可见,减毒鼠伤寒沙门氏菌能够引起宿主肠道菌群的代谢紊乱,本研究通过对小鼠粪样代谢组的研究能够更直观地得到宿主和菌群本身的代谢信息以及二者相互作用的共代谢信息.但是,粪样代谢组研究作为代谢组学研究中的一部分,其所能提供的代谢信息仍相对有限,未来还可以进一步开展减毒鼠伤寒沙门氏菌感染对小鼠不同脏器及各种体液代谢组的影响等方面的研究,以获取更多与减毒鼠伤寒沙门氏菌感染相关的宿主代谢信息. 3 结论 本研究同时使用 WIPM 和 Bruker 500 MHz NMR 对谱仪对同一批减毒鼠伤寒沙门氏菌感染小鼠的粪样进行检测,多变量数据分析结果均发现减毒鼠伤寒沙门氏菌感染能够引起小鼠粪样代谢物中多种氨基酸和尿嘧啶含量的上升以及胆汁酸、乳酸和丙酸含量的下降.这些变化的物质均说明减毒鼠伤寒沙门氏菌感染对小鼠宿主和肠道微生物的共代谢产生了一定的影响.以上研究为减毒鼠伤寒沙门氏菌的研究工作补充了基础的粪样代谢组信息.此外,本文首次开展了基于自主研发 NMR 波谱仪的代谢组学研究,该研究结果与利用极具权威的 Bruker NMR 波检仪检测所得到的结果高度一致.这点在某种程度上表明, WIPM NMR 波谱仪不仅能够满足科研人员对生物样品的 NMR检测需求,同时还能够确保检测数据在后续分析中的准确可信.该部分工作为基于自主研发 NMR波谱仪的代谢组学技术相关应用的开发奠定了基础,同时也为 WIPM NMR波谱仪的产业化提供可靠的科研依据. 致谢:感谢美国亚利桑那州立大学施一燊教授和武汉大学郭林教授对本实验所用减毒鼠伤寒沙门氏菌菌株的惠赠,同时感谢科技部谱仪项目(2011YQ12003503))的支持. ( 参考文献: ) ( [1] E verest P, K etley J , H ardy S , et a l. E valuation of s almonella typhimurium mutants i n a m odel o f e x perimental g astroenteritis[J]. Infect Imm u n, 1999,67(6):28 1 5一2821. ) ( [21 H ur J, L ee J H . I mmune responses to n e w vaccine candidates constructed b y a l i ve attenuated salmonella typhimuriumdelivery system expressing escherichia coli f4, f 5 , f6, f 41 a nd intimin adhesin antigens in a murine model[J]. J Vet Med Sci, 2011,73(10) : 1265 -1 273. ) ( [3] X in W, L i Y , M o H, et al. P s pa family fusion proteins delivered by a t tenuated s a lmonella e n terica serovar typhimuriumextend and enhance protection against streptococcus pneumoniae[J]. Infect Immun, 20 0 9,77(10):4518-4528. ) ( [4] Yang X L , L i u W C, Ya n g W W, et a l . Ora l imm u nization of mice with vacc i ne of attenuated salmonella typ h imuriumexpressing helicobacter pylori urease b subunit[J]. Biomed E nviron Sci, 2005, 18 (6):411-4 1 8 . ) ( [5] Rahman M M, Uyangaa E, Han Y W, et al. Oral administration of live attenuated salmonella enter i ca serovar typhimurium expressing c h icken in t erferon-alpha all e viates clinical signs cau s ed by r e spiratory infection with avian influen z a virus h9n2[J]. Vet Microbiol, 201 1 , 154(1-2): 1 4 0一151. ) ( [6] Cheng M ing(程明), He Jin-sheng(何金生), Fu Yu a n-hui(付远辉), et al. An t ibody responses induced b y mucosal DNAvaccine encoding t he codon-optimized F p r otein of human respiratory s y ncytial v irus (RSV) delivered w ith a ttenuated salmonella typhimurium(以减毒鼠伤寒沙门菌为载体的人呼吸道合胞病毒F蛋白DNA疫苗滴鼻免疫抗体分析)[]. Acta Microbiologica Sinica(微生物学报),2011,51(7):965-971. ) ( [7] Chen G, D a i Y, Chen J, et al. O r al delivery of the sj23lhd-gst antigen by salmonella typhimurium type i i i secretion systemprotects against schistosoma japonicum infection in mi c e[J]. PL o S Negl Trop Dis, 2011,5(9): e1313. ) ( [8] Y ang Y , Zhang Z, Ya n g J, et al. O r al v a ccination with ts87 DNA vaccine deli v ered by a t tenuated salmonella typhi m urium elicits a protective immune response against trichinella spiralis larval challenge[J]. Vaccine, 2010,28(15):2735-2742. ) ( [9] Qu D, Y u H, Wang S, e t al. In d uction of p r otective immunity by multiantigenic D N A vaccine del i vered in at t enuatedsalmonella typhimurium against toxoplasma gondii infection in mice[J]. Vet Parasitol, 2 009,166(3-4):220-227. ) ( [10] 0 ] T ang H, Wang Y. Metabonomics: A r evolution in progress[J]. Prog Biochem Biophys, 2006,33(5):401一417. ) ( [11 ] C ( hen L u (陈璐), Wang Yu-lan(王玉兰) . M e t abo l ite compo s ition of planarian ana l yzed by n uclear magnetic re s onance spectroscopy (涡虫代谢物组成的核磁共振分析)[J]. Chinese J Magn Reson(波谱学杂志 ) ,2013,30(4): 516-527. ) ( [12 ] Cevallos-C e vallos J M, D a nyluk M D, Reyes-De-Corcuera J I. Gc-ms b ased metabolomics for rapid simultaneous dete c tionof escherichia c oli o157:H7, salmonella t y phimurium, salmonella muenchen, and s a lmonella hartford in gro u nd beef an d chicken[J].J Food Sci, 2011,76(4): M238-M246. ) ( [13 ] A / ntunes L C M, Ar e na E T, Menendez A, e t a l . Impact of s almonella inf e ction on host hormone metabolism revealed by metabolomics[J]. Infect Immun, 2011,79(4):1759-1 7 69. ) ( [14 ! ] ] C lairmont C , L e e K C , P ik e J, et a l. Bio d istribution an d gen e tic sta b ility of the no v el ant i tumor age n t vnp2 0 009, agenetically modified strain of salmonella typhimurium[J]. J Infe c t Dis, 200 0 ,181(6):1996-2002. ) ( [15 ] Cl ( airmont C, Be r mudes D, Ittensohn M, et a l . Genetically modified salmonella typhimurium, vnp20009, a novel anticanceragent: Bio-distribution, genetic stability, selective tumor accumulation, and antitumor efficacy[J]. C a ncer Gene Ther, 1999, 6(6): S11-S12. ) ( [16] 5 ] N icholson J K, Connelly J, Lindon J C, et al. Metabonomics: A platform for studying drug toxicity and gene function[J]. Nat Rev Drug Discov, 2002,1(2):153-1 6 1. ) ( [17] Tian Yuan(田园), Tang Hui-ru(唐惠儒). Identification and structural determination of saccharides in rat feces( 大 鼠粪样中 几种糖类物质的结构确定)[J].Chinese J Magn Reson(波谱学杂志) , 2012,29(3):361-37 1 . ) ( [18] J olliffe I . Principal Component Analysis[M]. Wiley Online Library: 2005. ) ( [19 9 ] ]E riksson L, T rygg J , Wold S. CV-ANOVA for signi f icance testing of p l s a nd opls @ mode ls [J]. J Chemometr, 2008, 22(1 1 -12):594-600. ) ( [201 0 ] J iang L , H uang J, Wang Y, e t al. Metabonomic analy s is revea l s the ccl4-induced systems alterations for multiple rat o rgans[J]. J Proteome Res, 2012, 11(7) : 384 8 -3859. ) ( [21 ] Fan T W M, La n e AN. Structure-based profiling of metabolites and isotopomers by NMR[J]. Prog Nucl Mag Res Sp, 2008 , 52(2-3):69-117. ) ( [22] Wang Y, Tang H, Hol m es E, et al . B i och e mical charac t erization of rat in t est i ne devel o pment using high-resolution magic-angle-spinning 1h nmr spectroscopy and multivariate data analysis[J].J Proteome Res, 2005, 4(4) : 1 324一1329. ) ( [231 3 ] Marchesi J R, H o lmes E, Khan F, et al . R a pid and noninvasive metabonomic characterization o f inflammatory boweldisease[J]. J Proteome Res, 2007 , 6(2):546-5 5 1. ) ( [241 4 ] J ohnston S J , R i dge S A , Cassidy J, e t a l. Regulation of d ihydropyrimidine dehydrogenase in colore c tal cancer [ ]. ClinCancer Res, 1999,5(9):2 566-2570. ) ( 251 5 ] J ones B V , Begley M, Hi l l C, et al. Functional a n d comparative metagenomic analysis of bile salt hydrolase act i vity in th e human gut microbiome[J]. Proc Natl Acad Sci USA, 2008, 105(36):13 580一135 8 5. ) ( 26 5 1 ] Grubb B R . Avian cecum: Role of glucose and volati l e fatty acids in trans e pithelial ion t ransport[J]. Am J Physi o l-Gastr L, 1991 , 260(5): G703 - G710. ) ( [27] G ill S R , Pop M, DeBoy R T, et al. M et a genomic analysis of th e human distal gut m i crobiome[J]. Science, 2006,312(5778): ) ( 1 355一1359 ) [28]]]Bourriaud C, Robins R, Martin L, et al. Lactate is mainly fermented to butyrate by human intestinal microfloras butinter-individual variation is evident[J]. J Appl Microbiol, 2005,99(1): 201-212. Effects of Attenuated Salmonella Typhimurium Infection onFecal Metabonome in Mice comparison between WIPM and Bruker 500 MHz NMR spectrometers CHEN Lul2, SONG Kan13, WANG Yu-lanl* [1. Key Laboratory of Magnetic Resonance in Biological Systems, State Key Laboratory of Magnetic Resonance and Atomic and Molecular Physics, National Center for Magnetic Resonance in Wuhan (Wuhan Institute of Physics and Mathematics, Chinese Academy of Sciences), Wuhan 430071, China; 2. Center for Disease Prevention and Control of Changjiang River Administration of Navigational Affairs,Wuhan 430019, China; 3. University of Chinese Academy of Sciences, Beijing 100049, China] Abstract: Salmonella typhimurium iiss a intracellular, invasive, facultative anaerobicbacteria. With improved safety, attenuated salmonella typhimurium has been widely used indevelopment of vaccine for preventing viruses, bacteria, and parasites infections. However,limited research has been done so far on the biological effects of attenuated salmonellatyphimurium infection on animal hosts. In this work, we measured fecal metabonome ofmice infected with attenuated salmonella typhimurium using NMR spectroscopy. Two setsof NMR data were acquired with the same protocols on a Bruker 500 MHz NMRspectrometer and an in-house 500 MHz NMR spectrometer developed in Wuhan Instituteof Physics and Mathematics (WIPM). Multivariate data analysis methods were used toanalyzed the data. It was found that infection induced increases of certain amino acids anduracil in feces, and decreases of bile acids, lactate and propionate. Such metabolicalterations indicated that infection of attenuated salmonella typhimurium may causedisturbance of gut microbiotal communities. The results obtained from the home-developedspectrometer in WIPM showed high consistency with those obtained from Bruker 500 MHzNMR spectrometer, suggesting that the WIPM spectrometer is ready for metabonomicstudies. Key words: NMR, fecal metabonome, attenuated salmonella typhimurium, spectrometer ( *Corresponding author: WANG Yu-lan, T e l:+86-27-87197143, E-mail: yulan.wang@wipm.ac.cn. ) WIPM-I核磁共振波谱仪是我国首个进行商品化生产的高场谱仪,该产品具有以下特点: (1)采用基于网络的分布式架构设计,各模块可独立工作,通道数量可任意扩展,并可根据用户需求定制; (2)数字化接收机采用直接中频采样和数字滤波技术,消除了传统谱仪的镜像峰干扰问题; (3)基于 CAN 总线的控制器设计; (4)模块化设计,系统高度集成,结构简洁; (5)用户熟悉的 Windows 环境,控制软件 SpinStudio 支持图形和命令行操作方式,.实验数据可与 Varian, Bruker, Mestnova 转换; (6)提供高效的梯度匀场、搜索、3D自动匀场功能和业内最优秀的自动基线校正、自动相位校正算法; (7)集成了大量成熟的脉冲序列,并提供图形化的脉冲序列编辑器,用户可使用高级语言自行编写脉冲序列; (8)参数可自动优化,无需繁杂的实验参数设置 鼠伤寒沙门氏菌属于胞内侵袭性兼性厌氧菌,减毒后的鼠伤寒沙门氏菌安全性大大提高,已被广泛用于病毒、细菌及寄生虫疫苗的研制.但是目前还没有研究利用代谢组学方法来分析减毒鼠伤寒沙门氏菌感染对宿主和肠道菌群相互作用的影响.同时中国科学院武汉物理与数学研究所自主研发的 WIPM 500 MHz 核磁共振(NMR)波谱仪已经研制成功,但是该谱仪是否可以用于代谢组学研究尚未经过考证.本研究同时使用 WIPM 和 Bruker500 MHz 核磁共振波谱仪对同一批减毒鼠伤寒沙门氏菌感染小鼠的粪样进行检测,并结合多变量数据分析方法来研究减毒鼠伤寒沙门氏菌感染对小鼠粪样代谢组的影响.研究结果表明, WIPM NMR 波谱仪检测数据的分析结果与 Bruker NMR 波谱仪的实验结果具有高度的一致性,表明 WIPM 谱仪可以用于代谢组学研究.研究发现减毒鼠伤寒沙门氏菌感染能够引起小鼠粪样代谢物中多种氨基酸和尿嘧啶含量的上升,并伴有胆汁酸、乳酸和丙酸含量的下降.以上代谢物含量的改变表明,减毒鼠伤寒沙门氏菌感染使小鼠肠道微生物的代谢功能发生了紊乱.
确定










还剩13页未读,是否继续阅读?

产品配置单

武汉中科牛津波谱技术有限公司为您提供《小鼠粪样代谢产物中感染减毒鼠伤寒沙门氏菌含量检测方案(核磁共振)》,该方案主要用于粪便中感染减毒鼠伤寒沙门氏菌含量检测,参考标准--,《小鼠粪样代谢产物中感染减毒鼠伤寒沙门氏菌含量检测方案(核磁共振)》用到的仪器有中科牛津 WNMR-I 400-600MHz核磁共振波谱仪
推荐专场





相关方案
更多