推荐厂家
暂无
暂无

 400-629-8889
400-629-8889
 留言咨询
留言咨询
 400-860-5168转0230
留言咨询
400-860-5168转0230
留言咨询
 400-807-5250
留言咨询
400-807-5250
留言咨询

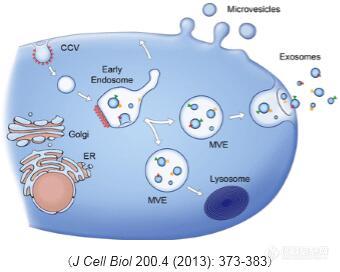
何为细胞外泌体? 外泌体最早发现于体外培养的绵羊红细胞上清液中,是细胞主动分泌的大小较为均一,直径为40~100纳米,密度1.10~1.18 g/ml的囊泡样小体。细胞外泌体携带多种蛋白质、mRNA、miRNA,参与细胞通讯、细胞迁移、血管新生和肿瘤细胞生长等过程并且有可能成为药物的天然载体,应用于临床治疗。 然而,测量技术手段的局限限制了外泌体在这些领域的研究进展。所以,在这篇文章中,作者总结了外泌体的纯化方法(离心法、过滤离心法、密度梯度离心法、免疫磁珠法以及色谱法),比较了现存各种外泌体测量技术(电子显微镜、动态光散射技术及纳米微粒追踪分析术)在外泌体尺寸和表征研究中的应用。原文点击——综述:细胞外泌体颗粒表征测量技术新进展
【背景】CIK是“Cytokine-Induced Killer Cells”的缩写,中文全称为“细胞因子诱导的杀伤细胞”。 CIK是单个核细胞在CD3单抗和多种细胞因子(包括IFN-g, IL-2等)的作用下培养获得的一群以CD3+CD56+细胞为主要效应细胞的异质细胞群, 其既具有T淋巴细胞强大的抗肿瘤活性,又具有NK细胞(自然杀伤细胞)的非MHC(主要组织相容性抗原)限制性肿瘤杀伤能力。CIK细胞具有杀瘤活性高、杀瘤谱广,对正常组织毒性低,体外可高度扩增等特点,是目前临床上广泛使用的过继性免疫治疗细胞。【培养原理】CIK培养用细胞因子和抗体:nCD3激发型单抗:T细胞活化的第一信号来自于T细胞表面的受体,即T细胞抗原受体(T cell antigen receptor, TCR)与APC提呈的抗原的特异性结合,也就是T细胞对抗原的特异性识别。TCR是由2条不同肽链构成的异二聚体,在T细胞表面,其与CD3分子通过非共价键结合,形成TCR/CD3复合体。TCR识别特异性抗原后会引起CD3和T细胞表面的辅助受体CD4或CD8分子的胞浆尾部聚集,进而激活与胞浆尾部相连的酪氨酸激酶(Lck, Fyn和ZAP-70等),促使CD3分子胞浆区的免疫受体酪氨酸活化基序(immunoreceptor tyrosine-based activation motif, ITAM)中的酪氨酸(Y)磷酸化。磷酸化的酪氨酸(pY)进一步磷酸化下游含酪氨酸的蛋白,从而引起激酶活化的级联反应(磷脂酰肌醇途径或MAP激酶途径等),最终通过激活转录因子,使其进入细胞核内,结合于调控T细胞增殖和活化的靶基因(如IL-2和IFN-g等),引起基因的表达和转录,T细胞因而由静止状态转为增殖和活化状态。由上可见,CD3分子在T细胞活化信号的转导中起着极其关键的作用。CD3激发型单抗与T细胞表面CD3分子特异性结合后,可引起CD3分子胞浆区ITAM基序中酪氨酸的磷酸化,进而导致T细胞增殖和活化的下游信号的激活,从而使T细胞增殖和活化。也就是说,CD3激发型单抗能够模拟抗原与TCR/CD3复合物的识别和激活过程,从而引起T细胞的增殖与活化,因此是CIK细胞培养中不可或缺的刺激因素。此外,CD3激发型单抗在选用时一定要注意克隆号。研究表明,仅克隆号为OKT-3的CD3激发型单抗可以刺激所有人的T细胞的增殖,而其它克隆号的CD3激发型单抗仅能刺激一部分人的T细胞。因此,在进行CIK培养时,最好选用OKT-3克隆,以保证每个患者的T细胞均能被激活。nIL-2 (白细胞介素-2)IL-2最初发现时被称为T细胞生长因子(T cell growth factor, TCGF),是引起T细胞增殖最重要的细胞因子。IL-2既是自分泌细胞因子,也是旁分泌细胞因子,其通过与T细胞表面的IL-2受体(IL-2R)的特异性结合而促使T细胞活化,并进入细胞分裂状态。此外,IL-2还可刺激NK细胞的生长并增强其杀伤能力。因此CIK细胞培养中须添加IL-2,以促进T细胞的增殖与活化。nIFN-g (干扰素-g)IFN-g 具有上调外周血淋巴细胞表面IL-2R表达的作用,因此会增强T细胞对IL-2促增殖反应的敏感度和强度。在诱导CIK细胞形成的过程中加入IFN- g ,可降低IL-2的用量。研究发现,IFN-g加入的顺序与CIK的细胞毒活性密切相关。先加入IFN- g,培养24后再加入IL-2,可明显提高CIK的细胞毒活性。nIL-1a(白细胞介素-1a)IL-1a也可以介导外周血淋巴细胞表面上调表达IL-2R。当IL-1a与IFN-g和激发型CD3单抗合用时,可以明显提高CIK 的细胞毒作用。【细胞制备】1.外周血单个核细胞的采集1.1用血细胞分离机采集患者自身的外周血单个核细胞50-100mL;1.2淋巴细胞分离液密度梯度离心法进一步纯化单个核细胞(PBMC);1.3无血清培养液洗涤2次,获得纯度在90%以上的PBMC。2.CIK细胞的培养及鉴定2.1将PBMC按1-2 x 106/ml的浓度悬浮于无血清培养液中,加入1,000 U/ml 的重组人IFN-g,37oC,5%CO2培养箱中培养;2.224h 后加入50ng/ml 的CD3 单克隆抗体和300 U/ml 的重组人IL-2,刺激CIK 细胞的生长和增殖;注:此时也可同时加入100 U/ml的重组人IL-1a。2.3每3天半量换液或扩瓶一次,并补加重组人IL-2 300 U/ml;2.4在培养的第14d,收获CIK细胞。2.5CIK细胞质控:2.9.1台盼蓝染色检测:活细胞应在80%以上;2.9.2流式细胞仪检测细胞表面CD3、CD8、CD56等分子的表达:CD3+CD56+细胞的比例应在20%以上。2.9.3细胞杀伤实验:以CIK细胞为效应细胞,以肿瘤细胞(可为原代肿瘤细胞或肿瘤细胞株)为靶细胞,将效应细胞与靶细胞按10 : 1(数目比) 的比例加入96 孔U 型板中,每孔含靶细胞1 x 104个,终体积为200 ml,设3个复孔。培养4h,然后取培养上清,用乳酸脱氢酶(LDH) 试剂盒检测效应细胞对靶细胞的杀伤率。2.9.4收获细胞前,取少量培养物进行细菌、真菌培养,并检测支原体、衣原体,及内毒素(标准:病原学检测阴性,内毒素5 Eu)。【步骤简图】http://img.dxycdn.com/trademd/upload/userfiles/image/2013/04/B1366873006_small.jpg 【推荐试剂】http://img.dxycdn.com/trademd/upload/userfiles/image/2013/04/B1366873008_small.jpg 注:Animal Free意为无动物成分。无动物成分的重组细胞因子在生产过程中不会有任何动物源性物质,尤其是牛蛋白的混入,使得最终获得的重组人蛋白中不含任何动物成分。这样可避免动物病原体(如疯牛病,克雅氏病等)的污染及外源蛋白引起的机体异种排斥和过敏反应,因此细胞治疗的体外细胞培养过程中最好使用无动物成分的重组细胞因子。【其它相关试剂】 http://img.dxycdn.com/trademd/upload/userfiles/image/2013/04/B1366873009_small.jpg【参考文献】 Li R, Wang C, et al. Autologous cytokine-induced killer cell immunotherapy in lung cancer: a phase II clinical study. Cancer Immunol Immunother. 2012; 61:2125-2133
我们专题主要是研究胶原纤维,费伦有提过胶原纤维存在红外光传输的特征波段,不过在国内外研究极少提到有关细胞与细胞外基质的光传输,大都是提到有关化学反应的过程,现在是想找在胶原纤维的UV与IR光谱里那一个波峰,透过纤连蛋白(fibronectin)传输光讯号到细胞上的受体,之前是有找到有关细胞不含胞器(只剩肌动蛋白丝actin、整键蛋白integrin)也能移动,所以我们就假设细胞的移动可能是胶原纤维所操控,讲得有点多了,因为就只差这一点专题就完成了,可以请专家提出一些意见吗?谢谢[IMG]http://www.cella.cn/book/10/images/image006.jpg[/IMG]







 我要推广仪器
我要推广仪器
 下载APP
下载APP