推荐厂家
暂无
暂无



 400-831-3689
400-831-3689
 留言咨询
留言咨询
 400-876-8980
留言咨询
400-876-8980
留言咨询
 400-860-5168转2077
留言咨询
400-860-5168转2077
留言咨询



农药对食用菌菌丝生长的影响 食用菌的农药残留问题受到更多的社会关注,目前造成农药残留超标的的主要原因有一是施药技术不到位,设备落后,施用农药过程中操作不规范,污染环境和作物;二是选用农药的种类、剂量、安全间隔施药不合理,滥用高毒、高残留农药;三是不在安全间隔期收获农作物,从而威胁食用菌产品质量安全。本文研究了农药对食用菌菌丝生长影响,探索了安全使用农药的方法,对食用菌安全生产有一定的指导作用。1 材料和方法1.1 试验材料1.1.1 农药种类 德国拜耳作物科学有限公司的2.5%溴氰菊酯乳油 江苏扬农化工集团有限公司的10%氯氰菊酯乳油 山东圣鹏农药有限公司的10%联苯菊酯乳油 浙江威尔达化工有限公司的20%甲氰菊酯乳油 江苏苏州佳辉化工有限公司的480克/升毒死蜱乳油 南通江山农药化工股份有限公司的77.5%敌敌畏乳油 江苏龙灯化学有限公司的70%可湿性粉剂 江苏扬农化工有限公司的70%甲基托布津可湿性粉剂和50%多菌灵可湿 性粉剂1.1.2 菌种试验食用菌有5种,分别为真姬菇、黑木耳、双孢蘑菇、平菇、白灵菇,均为福建省主栽食用菌的品种,菌种由*******菌物研究中心提供。1.2 方法1.2.1 PDA培养基的配制去皮切块处理马铃薯,称取200 g,加蒸馏水1L煮沸,滤去土豆残渣,再加20 g葡萄糖和15~20 g琼脂煮沸,趁热充分溶解后纱布过滤定容,分装三角瓶,冷却后贮存备用。1.2.2未灭菌的农药对五种食用菌菌丝的影响农药浓度配置:农药对真姬菇、黑木耳、双孢蘑菇、平菇、白灵菇生长影响的试验中,每种农药设5个浓度处理(表3-1),共21个处理,包括一个空白对照。每个处理水平设3个重复。所用的农药在实验室预先配好母液,存放备用。农药对真姬菇、黑木耳、双孢蘑菇、平菇、白灵菇的生长影响:将农药(农药在高温下会分解,故而不高温灭菌)经过滤菌器后(粉剂不过滤)添加至已灭菌的PDA培养基的三角瓶中,按照表2的处理水平配置成含有农药的培养基后倒平板,冷却待用。用直径为1 cm的无菌打孔器打孔母种的培养基,挑取统一生长情况的母种块接种于上述农药浓度水平处理平板中,以纯PDA平板作为空白对照组,置于23-25℃下培养,待空白对照组菌丝生长至平板的三分之二,结束实验观察,测量各个处理水平菌落半径的大小。[img=,622,434]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301423_01_2903169_3.png[/img]2 结果与分析2.1农药对五种食用菌菌丝的影响2.1.1 九种农药对真姬菇的菌丝生长影响[img=,660,511]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301423_02_2903169_3.png[/img]从表2可知,敌敌畏与溴氰菊酯对真姬菇的菌丝有抑制作用。高浓度和低浓度有极显著差异,在浓度为0.2mg/kg时候,菌丝生长速度和0.1mg/kg、0.4mg/kg相比有显著差异。在0.1 mg/kg~0.4 mg/kg真姬菇生长速度增加,在浓度0.4mg/kg时,生长速度达到最大,在0.6 mg/kg~1.0 mg/kg真姬菇生长速度减小。百菌清在0~0.4mg/kg浓度范围内,菌丝生度速度显著差异随着浓度繁升高,真姬菇生长速率不断减慢。甲基托布津添的空白组与高浓度有显著差异,其他浓度之间在5%水平无显著差异,即在0.1 mg/kg~1 mg/kg之间,真姬菇菌丝生长速度无明显差异。氯氰菊酯在0.1 mg/kg~1 mg/kg之间会促进真姬菇生长,甲氰菊酯在0.4mg/kg浓度与其它处理有极显著差异(除1.0 mg/kg的水平浓度外)。联苯菊酯浓度为0.1mg/kg与空白组、中、高浓度有极显著差异,且菌丝生长速度最大,随后生长速率逐渐降低。毒死蜱的添加浓度不断增加时,真姬菇的菌丝先增加后减小,在添加浓度为0.01mg/kg与0.1mg/kg时,生长情况有极显著差异,浓度为0.05mg/kg时,真姬菇菌丝生长最快。2.1.2 九种农药对黑木耳的菌丝生长影响 [img=,624,474]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301424_01_2903169_3.png[/img]由表3可知,敌敌畏、溴氰菊酯以及联苯菊酯对黑木耳的菌丝有抑制作用,联苯菊酯在浓度高于0.1mg/kg时会抑制黑木耳生长。多菌灵在浓度高于0.2mg/kg时会抑制黑木耳生长。百菌清在浓度为0.2mg/kg与1mg/kg水平浓度有极显著差异,与其他浓度无极显著差异。甲基托布津在浓度为0.2mg/kg与空白组无极显著差异,与其他处理的浓度有极显著差异,甲基托布津浓度为0.2mg/kg时,黑木耳菌丝生长最快。当氯氰菊酯的添加浓度由低升高时,黑木耳的菌丝生长先增大后减小,当浓度是0.2mg/kg时,与其这处理的菌丝生长速度有极显著差异。不同浓度的甲氰菊酯处理,结果有极显著差异,甲氰菊酯浓度是0.2mg/kg,黑木耳菌丝生长最快。不同浓度的毒死蜱对菌丝生长情况不相同,当浓度为0.2mg/kg时,菌丝生长速率达到最大,同时和其他浓度的生长有极显著差异。2.1.3九种农药对双孢蘑菇的菌丝生长影响 [img=,636,476]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301424_02_2903169_3.png[/img]从表4可以发现,敌敌畏与溴氰菊酯抑制双孢蘑菇的菌丝生长。多菌灵在添加浓度为0.6mg/kg时候,菌丝生长最快。百菌清六个处理培养双孢蘑菇,所有处理水平之间无极显著差异。百菌清的在0.2mg/kg时,双孢蘑菇的菌丝生长速度最快。甲基托布津、甲氰菊酯的添加浓度为0.2mg/kg与添加浓度为0.4mg/kg的处理组分的结果之间有极显著差异,说明甲基托布津、甲氰菊酯的添加浓度是0.2mg/kg时,双孢蘑菇的菌丝生长率达到最大。在氯氰菊酯添加浓度不断增变大的过程中,双孢蘑菇生长先变大后减小,在浓度达到0.4 mg/kg时,生长速率最快,同时该组分处理与其他组分处理呈极显著差异。联苯菊酯浓度为0.2mg/kg~0.4 mg/kg与空白组和最高浓度的结果有极显著差异,在0.2mg/kg浓度水平,双孢蘑菇生长达到最大。毒死蜱的浓度为0.01mg/kg时,双孢蘑菇生长达到最大,与空白组无极显著差异性,与其他组分处理有极显著差异。2.1.4九种农药对平菇的菌丝生长影响 [img=,628,459]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301424_03_2903169_3.png[/img] 由表5可以看出,敌敌畏与溴氰菊酯对平菇的菌丝有抑制作用。对平菇在多菌灵的六个浓度中生长速度进行显著分析,0.2mg/kg~0.4 mg/kg的处理水平对空白组分、0.1 mg/kg的处理组份有极显著差异,说明在浓度0.2mg/kg~0.4 mg/kg时候,菌丝生长速度达到最大。百菌清在0.2mg/kg处理组份与其他组份有极显著差异,当百菌清添加浓度由低至高时,平菇生长速度先增加最大后减小。甲基托布津在浓度为0.2mg/kg~0.4 mg/kg时的结果与其他处理水平有极显著差异,在甲基托布津在浓度为0.2mg/kg时,平菇生长最快。随着氯氰菊酯的浓度不断变大,平菇的生长速度不断减小,中、低浓度组份生长情况与高浓度的有极显著差异。低浓度处理组份的甲氰菊酯对平菇生长无明显影响,高浓度处理组份抑制平菇菌丝生长。在联苯菊酯的处理组份为0.2mg/kg~0.4 mg/kg与空白组、高浓度处理组份有极显著差异,故而在0.2mg/kg浓度时,平菇生长达到最大。在毒死蜱处理水平不断增加时,菌丝生长先增大后减小,在0.01mg/kg~0.05mg/kg生长到最大,且与其他组份浓度生长情况有极显著差异。2.1.5九种农药对白灵菇的菌丝生长影响 [img=,636,441]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301425_01_2903169_3.png[/img] 由表6可知:敌敌畏、溴氰菊酯以及联苯菊酯对白灵菇的菌丝有抑制作用。当联苯菊酯的添加浓度大于0.1mg/kg时对白灵菇有抑制作用。多菌灵在浓度为0.2mg/kg时(除空白组外)与其他水平处理有极显著差异,菌丝生长速度先增大都减小。当甲基托布津、百菌清添加浓度为0.4mg/kg时,试验结果与其他水平处理有极显著差异,即菌丝生长速度先增大都减小。从表可以看到低浓度对白灵菇生长情况有促进作用,在浓度0.1mg/kg之后生长速度减慢。在甲氰菊酯在0.2mg/kg时,与其他水平处理有极显著差异,说明在0.2mg/kg浓度白灵菇生长速度达到最大值,生长趋势是先增大后减小。当毒死蜱的浓度有小变大的过程中,白灵菇的生长速度先变快后减慢,当浓度为0.5mg/kg时,生长速度与其他水平处理有极显著差异。3小结与讨论采用PDA平板法,研究了9种农药对5种食用菌菌丝生长的影响。在实验操作过程中由于农药会在高温中消解,故农药直接加入PDA培养基,具有强烈的毒性和气味,可以清晰观察到添加的农药种类的不同时,对五种食用菌的生长菌丝的生长影响明显不同。当PDA培养基中所添加的农药浓度不断增加的过程中,部分菌丝生长所受到影响也越来越明显实验结果表明敌敌畏以及溴氰菊酯对菌丝的抑制作用最大,敌敌畏对真姬菇、黑木耳、双孢蘑菇、平菇、白灵菇这五类食用菌的菌丝生长抑制率达到100%,联苯菊酯使得平菇、双孢蘑菇的生长减慢,当联苯菊酯浓度大于0.5mg/kg时,黑木耳和白灵菇不生长。毒死蜱的浓度增加时,黑木耳、平菇、双孢蘑菇的菌丝生长逐渐减慢。对照空白组分,当甲基托布津的浓度不断变大时,白灵菇、黑木耳、平菇、双孢蘑菇的生长趋势均是先增大后减小,同时对真姬菇菌丝生长影响不大,所以在真姬菇菌丝生长过程中,可以使用该农药。多菌灵、氯氰菊酯对平菇、黑木耳生长有抑制作用。而百菌清对真姬菇和白灵菇生长有抑制作用,故而在食用菌菌丝生长过程中慎用对应的农药。
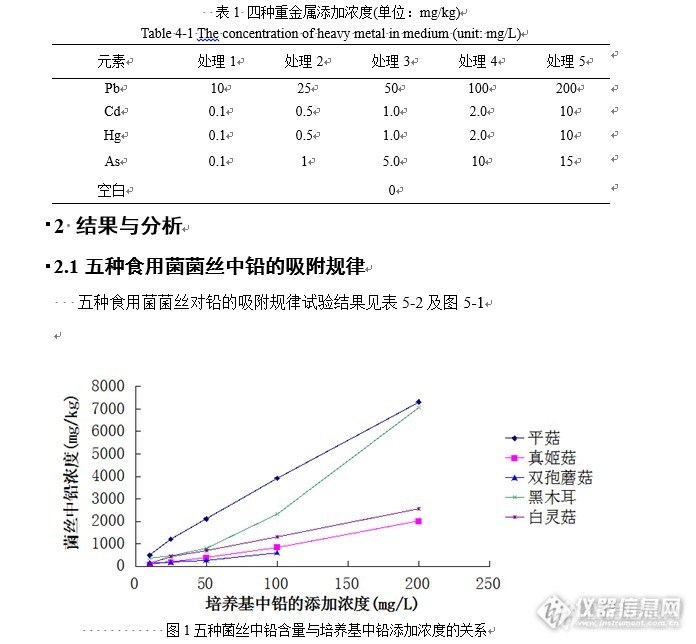
[b]食用菌菌丝对铅、镉、汞、砷的吸收规律[/b]重金属会富集于食用菌的子实体内。故此,人们对食用菌吸附重金属的能力越来越关注,食用菌吸附重金属的能力大小是由食用菌生长环境以及其本身相关的生物特性决定。研究食用菌菌丝吸附重金属的能力,对降低食用菌产品的重金属含量奠定了一定的基础。[b]1材料与方法1.1 试验材料1.1.1重金属的种类[/b] 硝酸铅(PbNO[sub]3[/sub]):1x500 g 国药集团化学试剂有限公司 氯化隔(CdCl[sub]2[/sub]):1x100 g 国药集团化学试剂有限公司 氯化汞(HgCl[sub]2[/sub]):1x250 g 国药集团化学试剂有限公司 三氧化二砷(As[sub]2[/sub]O[sub]3[/sub]):1x500 g 国药集团化学试剂有限公司[b]1.1.2菌种[/b]试验食用菌有6种,分别为真姬菇、黑木耳、双孢蘑菇、平菇、白灵菇,这些菌种是福建省主栽食用菌的品种,取自于******菌种研究中心。[b]1.1.3仪器[/b] AA-6300C 岛津石墨炉[url=https://insevent.instrument.com.cn/t/Wp][color=#3333ff]原子吸收[/color][/url]分光光度计 AA-7000 岛津原子荧光分光光度计[b]1.2 方法[/b]重金属药液浓度配置:铅、镉、汞、砷对真姬菇、黑木耳、双孢蘑菇、平菇、白灵菇的菌丝吸附规律的研究试验中,每种重金属设5个浓度处理(表5-1),以不添加任何重金属为空白对照。每个处理设3个重复,每个重复20瓶三角瓶。所用的重金属药液是预先配好的母液。铅配成20 g/L离子浓度的母液,镉、汞配成2 g/L离子浓度的母液,砷配成1.5 g/L离子浓度的母液,存放备用。先将配置好的液体培养基分装至250mL的三角锥形瓶中,每瓶150mL,再将重金属母液分别添加至各个瓶液体培养基中,于灭菌锅中高温灭菌。取出待冷却后后,在无菌的条件下分别加入3~4块母种块(取已生长好的菌种培养皿,用直径1cm打孔器打出的菌种块),再将三角瓶置于24℃、120r/min的摇床中,均匀振荡培养15-~20d,取出过滤菌丝球,检测菌丝中铅、镉采用石墨炉原子分光光度法,测定汞和砷采用原子荧光光度法测量。[img=,690,638]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301539_01_2903169_3.png[/img][img=,452,633]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301540_01_2903169_3.png[/img]注:富集系数=菌丝中重金属含量/培养基中重金属浓度,下同。Note:Enrichment coefficient=heavy metalscontent of fruit body/concentration of compost.Here in after注:本章节所有生长影响图单位(mg/L)与液体培养基的体积相对应[align=left] 从表5-2中可知:不同食用菌对铅的吸附能力不同,当铅浓度较低时(0~10 mg/kg)5种食用菌菌丝对铅的富集系数较大,双孢蘑菇和平菇的富集系数最大,分别为59.79和50.62;随着培养基中的铅添加浓度增大,5种食用菌菌丝的富集系数逐渐减小。当培养基中的铅添加浓度为0~10 mg/kg时,5种食用菌对铅的吸附能力大小为:双孢蘑菇、平菇>黑木耳>白灵菇>真姬菇;当培养基中的 铅添加浓度为25mg/kg~200mg/kg时,5种食用菌菌丝吸附重金属的能力大小为:平菇>黑木耳>白灵菇>真姬菇>双孢蘑菇。[/align][align=left] 培养基中铅的添加量(x)与菌丝中重金属含量(y)之间的关系用DPS分析后建立罗杰斯特曲线数学模型,通式即为:y=c/(1+e[sup]a-bx[/sup])。结合图5-1与表5-2发现,5种食用菌菌丝中铅的含量和培养基添加的铅浓度具有良好的拟合效果,R[sup]2[/sup]都在0.99以上。从图5-1可以发现当培养基中铅的添加浓度达到200mg/kg时,五种食用菌菌丝对铅离子的吸附量已近趋近饱和状态。[/align][align=left][img=,668,389]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301542_01_2903169_3.png[/img][/align][align=left][img=,495,618]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301542_03_2903169_3.png[/img][/align][align=left][/align][align=left][/align][align=left]由表5-3得出不同浓度处理下五种食用菌,菌丝所吸附的镉含量。当培养基中镉的添加浓度不同时候,双孢蘑菇和平菇菌丝对镉的富集系数不断增加,真姬菇和白灵菇菌丝对镉的富集系数先增加后较小,而黑木耳菌丝在镉添加浓度为1mg/kg之后几乎不变。当镉的添加浓度为2mg/kg时,双孢蘑菇的菌丝富集系数达到最大是319.7。当添加浓度为0.5mg/kg时,平菇菌丝对镉的富集系数达到最大为520.6。[/align][align=left][/align][align=left]当浓度为0.1mg/kg和2mg/kg时,对培养基中镉的吸附能力大小比较是:平菇>双孢蘑菇>黑木耳>白灵菇>真姬菇;当浓度为0.5mg/kg时,5种食用菌菌丝吸附能力比较是:平菇>真姬菇>双孢蘑菇>黑木耳>白灵菇。[/align][align=left][/align][align=left] 结合图5-2与表5-3发现,5种食用菌菌丝中镉的含量以及培养基添加的镉浓度相关性的拟合度不错,R[sup]2[/sup]都在0.99以上(只有真姬菇和平菇的罗杰斯特曲线相关系数在0.9以上)。通过分析可以得到用这种数学罗杰斯特曲线数学模型可以说明培养基中重金属与菌丝中镉含量具体的相关关系。从图5-2可以发现当培养基中镉的添加浓度达到10mg/kg时,五种食用菌菌丝对铅离子的吸附量已近接近饱和状态,而双孢蘑菇以及平菇在镉的添加浓度为2mg/kg以后,菌丝不生长,说明吸附能力已经达到饱和的状态。[/align][align=left][/align][align=left][/align][align=center]表5-4 不同浓度处理下五种食用菌菌丝中汞的含量及罗杰斯特方程[/align][align=left][/align][align=center]Table 5-4 The concentrations of Mercury in five mushrooms treated byheavy metals and Roger Lancaster equation[/align][align=left][/align][align=left][img=,660,391]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301544_01_2903169_3.png[/img][/align][align=left][img=,527,623]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301545_02_2903169_3.png[/img][/align][align=left]由表5-4得出不同浓度处理下五种食用菌,菌丝所吸附的汞含量。当培养基中汞的添加浓度增加时,平菇和双孢蘑菇的菌丝对汞的富集系数不断减小,而黑木耳、真姬菇菌和白灵菇的菌丝丝对汞的富集系数先增加后减小。真姬菇菌丝富集汞的系数在汞的添加浓度为0.5mg/kg时达到最大是610.3,真姬菇和白灵菇菌丝富集汞的系数分别在1mg/kg和2mg/kg时达到最大为100.5和159.7。五种食用菌对汞的吸附能力不相同。但总体上来说,培养基中添加不同汞离子浓度,真姬菇菌丝相对其他四种菇菌丝(双孢蘑菇、平菇、黑木耳、白灵菇)对汞的吸附能力强。[/align][align=left] 结合图5-3与表5-4发现,培养基添加的汞浓度与5种食用菌菌丝中汞的含量的数学罗杰斯特曲线相关系数R[sup]2[/sup]都在0.9以上,通过分析可以知道用该数学模型可以说明培养基中重金属与菌丝中汞含量的相关关系。从图5-2可以发现当培养基中汞的添加浓度达到10mg/kg时,五种食用菌菌丝对铅离子的吸附量已近趋近饱和状态,而双孢蘑菇以及白灵菇在添加浓度为2mg/kg以后,菌丝生长缓慢,说明对汞的吸附能力已经达到饱和的状态。[/align][align=left][img=,681,454]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301547_01_2903169_3.png[/img][/align][align=center]表5-5 不同浓度处理下五种食用菌菌丝中砷的含量及罗杰斯特方程[/align][align=center]Table 5-5 The concentrations of Arsenic in five mushrooms treated by heavy metalsand Roger Lancaster equation[/align][align=center][img=,519,616]http://ng1.17img.cn/bbsfiles/images/2017/09/201709301549_01_2903169_3.png[/img][/align][align=center][/align][align=center][/align][align=center][/align][align=left]由表5-5得出不同浓度处理下五种食用菌,菌丝所吸附的砷含量。当添加砷的浓度不同时,五种食用菌对砷的吸附能力不相同。另外当砷的添加浓度不断增加时,五种食用菌菌丝对砷的富集系数都随之减小。当砷的添加浓度为0.1mg/kg和1mg/kg时,五种食用菌对砷吸附能力大小比较是:双孢蘑菇>黑木耳>真姬菇>白灵菇>平菇;当浓度为5mg/kg和10mg/kg时,5种食用菌菌丝吸附能力比较分别是:真姬菇>黑木耳>白灵菇>平菇=双孢蘑菇、黑木耳>白灵菇>真姬菇>平菇,双孢蘑菇。[/align][align=center][/align][align=left]结合图5-4与表5-5发现,建立添加的砷离子浓度与5种食用菌菌丝吸附砷的含量的罗杰斯特方程。结果分析可以得出,该方程曲线的相关性均达到0.99以上,说明罗杰斯特方程可以较好说明添加的砷离子浓度与5种食用菌菌丝吸附砷的含量关系的拟合程度。[/align][align=center][/align][b]3 小结与讨论[/b][align=center][/align][align=left]在液体发酵培养基中添加不同重金属离子的浓度与菌丝中相对应的重金属含量作图(图5-1~5-4),从这几个图可以得出五种食用菌的菌丝对重金属残留吸附量与培养基中重金属的添加浓度的相关性,图中大多数表现为“S”型曲线的关系。当添加在培养基中的重金属的浓度不断增加时候,不同的菌丝对重金属的吸收大多数有小变化到大,最后趋于某个上限值。对曲线方程y=c/(1+e[sup]a-bx[/sup])的数学进行可以得到,当x值为0时候,y值为c/(1+e[sup]a[/sup]),而当x趋近无穷大时,y值也无限接近于c,即培养基中重金属量的添加浓度达到最大时,食用菌菌丝对重金属的吸附趋于一个极限值,即最大可能吸附量(只是一个纯理论值,在实际生产中要考虑食用菌菌丝生长环境的重金属污染及抑制作用)。[/align][align=center][/align][align=center][/align][align=center][/align][align=center][/align]
加热蛋白溶菌酶能杀灭诺如病毒日本东京海洋大学的一个研究小组日前宣布,在实验中发现,加热处理鸡蛋蛋白含有的溶菌酶,能灭活诺如病毒。这是由于溶菌酶能破坏包裹诺如病毒基因的外壳。诺如病毒会引发急性肠胃炎和食物中毒。这种病毒具有强大的感染力,只要有10至100个病毒体进入人体,就会导致感染,目前还没有有效的抗病毒剂。研究小组利用实验鼠的诺如病毒替代人类诺如病毒进行了实验。他们将蛋白中含有的溶菌酶在100摄氏度下加热40分钟,使其变性。接下来,将含有1%加热处理过的溶菌酶的溶液与实验鼠诺如病毒混合在一起,并观察了1分钟之后的变化。溶菌酶是蛋白等含有的一种能水解致病菌中黏多糖的碱性酶。研究人员发现,诺如病毒基因量大幅减少,以致无法检出,并观察到病毒体出现膨胀。他们认为这是由于包裹病毒基因的外壳被破坏导致的。研究人员指出,实验鼠诺如病毒和人类诺如病毒从遗传学上来看非常类似,所以这种加热变性处理的蛋白溶菌酶对人类诺如病毒应该也有效果。他们希望将其制成消毒喷雾剂,在下一年度达到实用化。










 我要推广仪器
我要推广仪器
 下载APP
下载APP