本文译自Werner等科学家在美国化学学会会刊(ACS)上发表的一篇综述,比较详实系统介绍了超高分辨率显微技术在神经科学上的最新应用进展。我们在此文基础上进行了编译整理。因文章较长,我们将分期陆续介绍。本期介绍第三部分。
 《Super-resolving Microscopy in Neuroscience》
《Super-resolving Microscopy in Neuroscience》

荧光成像技术在我们理解神经系统中起着关键作用。各种超分辨率显微镜方法和专用荧光探针的出现使得能够以迄今为止无与伦比的分辨率直接洞察细胞亚区室中的神经元结构和蛋白质排列。
神经元的超分辨率可视化技术揭示了对细胞骨架组成、分布、运动性和膜蛋白信号传导、突触下结构和功能以及神经元-胶质细胞相互作用的新认识。自身免疫性和神经退行性疾病模型中明确的分子靶点为使用新型和创新的成像方法深入研究疾病病理生理学提供了极好的起点。超分辨率显微镜在人脑样本和临床生物标志物检测中的应用仍处于起步阶段,但为神经学和神经科学的转化研究提供了新的机会。在这篇综述中,作者描述了超分辨显微镜在过去的二十年里是如何提高我们对神经元和大脑功能和功能障碍的理解的。
在这一节中,作者将介绍SRM最近获得的关于细胞骨架的组成和动力学、突触前和突触后区室中对神经传递保真度至关重要的分子组装以及塑造神经元功能的星形胶质细胞结构的调节和构建的数据。
5.1. 细胞骨架
神经元的极化性质以及树突和轴突的长度需要结构化和功能性支架来支持稳定性、适应性可塑性和货物运输,这是神经元存活和信号传导所不可或缺的。因此,神经元细胞骨架的结构布局在过去几十年中吸引了神经科学家的注意,并在其他地方进行了详细综述。20世纪70年代的EM研究揭示了三种主要类型的细丝组成神经元细胞骨架:微管大小约为20−30 nm,神经丝直径为10nm,肌动蛋白丝大小为5− 10 nm。微管是由异二聚体组装的圆柱体,异二聚体在GTP依赖性组装过程中结合α和β微管蛋白单体,导致形成定义一个微管单位的13条原丝之一。轴突微管组织成束,根据其相对于神经元胞体的位置显示不同的方向。它们极化为快速生长的正端和缓慢生长的负端。
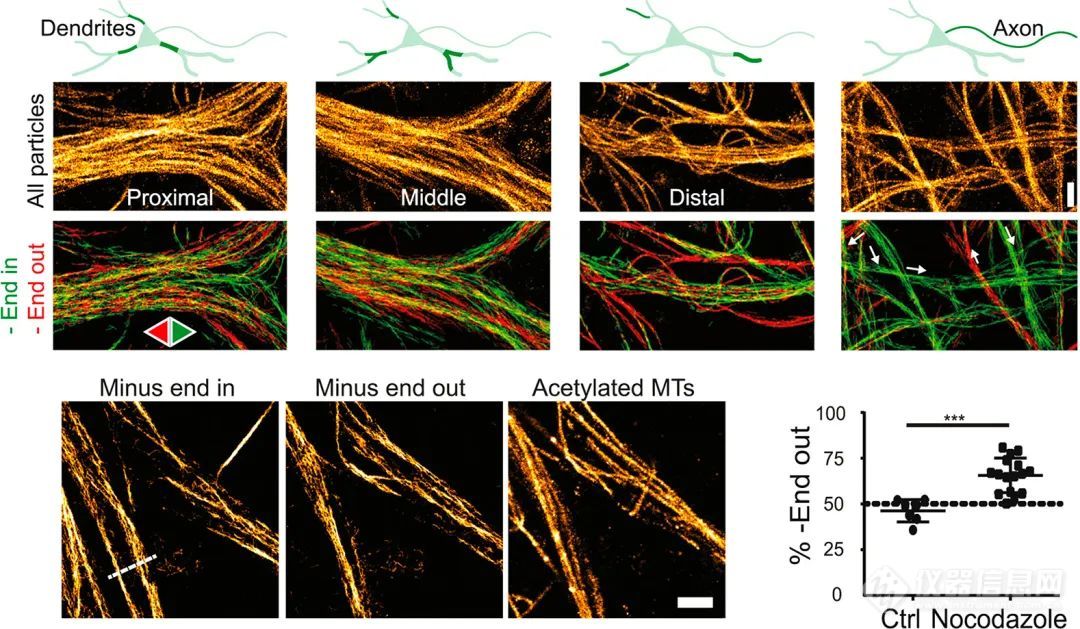
STED显微镜揭示了快速生长的末端在钙依赖过程中锚定在肌动蛋白皮质上。在发育中的神经元中,用dSTORM测量补充活细胞成像的实验证明了神经元极性和轴突规格,表明TRIM46驱动方向一致的平行微管束的形成,而树突微管的特征是混合极性。motor-PAINT的纳米跟踪显示,稳定的乙酰化微管显示负端向外的方向,而相反的方向在动态的酪氨酸化微管中占主导地位(图7)。由于密集的重叠定位,在神经束中密集排列的微管(例如轴突起始段)的SMLM具有挑战性。这个问题可以用两种实验方法来解决:第一,设计更小的标记探针,如微管蛋白纳米体,这有助于更详细地观察神经元微管。第二,一种去聚集超分辨率方法,如ExM,可用于显示索马和树突中的微管亚群。
图7.Motor-PAINT显示微管的方向
上排:源自跟踪原代大鼠神经元中荧光标记的运动蛋白的所有定位的总和(DIV 16 - 17)。下面一行:轨迹方向的颜色映射。比例尺= 1 μ m。左下角:乙酰化微管倾向于负端向外取向。右下角:抗诺考达唑处理的稳定微管显示普遍的负末端向外取向。显示平均值和标准差,t检验,*** p〈0.001。比例尺= 1 μ m。
神经丝是在轴突中形成大范围平行网络的杂聚物,提供轴突直径的稳定性和调节,从而提供传导速度。其组成包括低、中、高分子量神经丝、连接蛋白和外周蛋白的三联体。它们的自组装从平行异二聚体的形成开始,然后半交错组合成反平行四聚体。最后,八个四聚体横向聚集成一个单位长度的神经丝,该神经丝进一步伸长并径向压缩成最终的神经丝外观。在丝之间的界面上形成了3−5 nm大小的交叉桥,这已经用EM观察到,但对它们的功能及其与神经丝的分子相互作用的理解仍然缺失。这里,ExM结合SMLM的引入或DNA-PAINT的应用可能有助于阐明密集的神经丝中的这种相互作用。神经丝动力学已经通过光转换和光激活SRM实验进行了研究,显示了端到端的退火和切断过程。
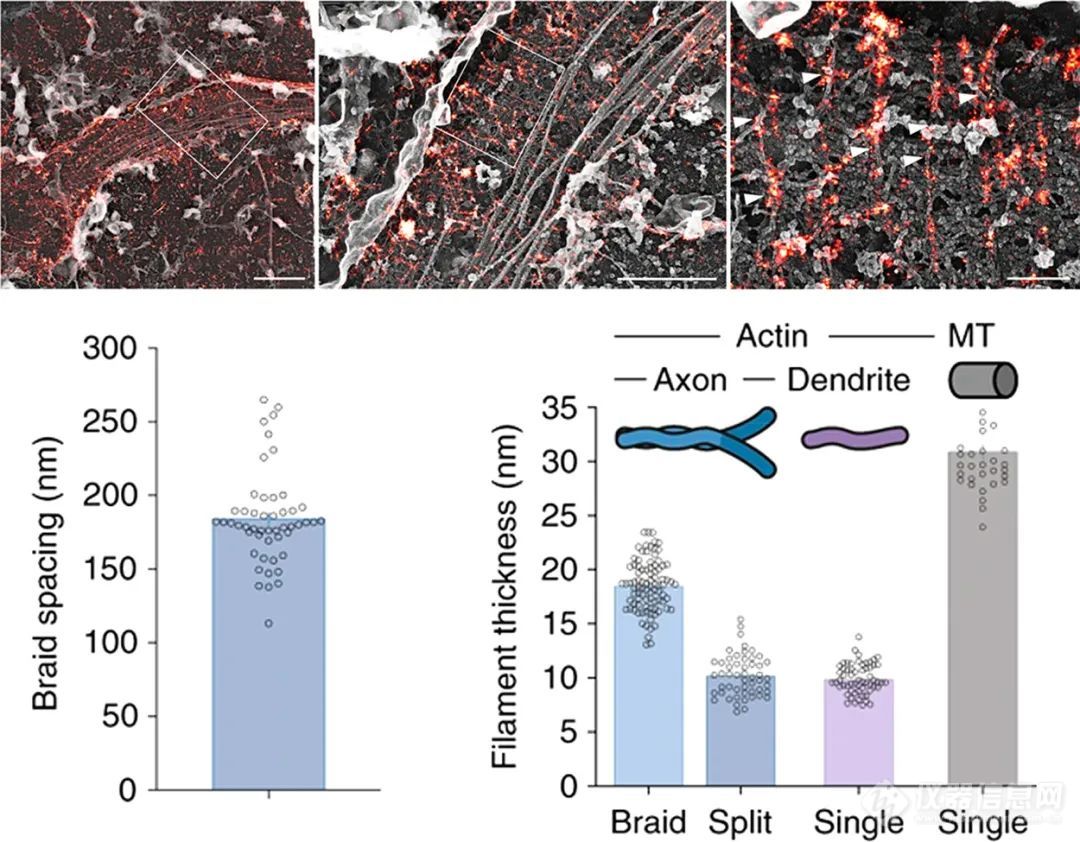
肌动蛋白最初被认为在轴质中形成斑点状膜下包被,并与一组更集中的短肌动蛋白丝结合。在原代神经元和脑切片中使用鬼笔环肽Alexa Fluor 647进行STORM记录,揭示了轴突肌动蛋白的新组织原则。这些实验揭示了轴突中存在表现出190 nm稳健重复间隔的环状肌动蛋白环,并进一步表征了轴突中β II血影蛋白和钠通道具有相似尺寸的周期性条带,而树突隔室显示出更长的肌动蛋白组织。此外,尽管通过STORM成像也在树突中显示了这种模式的普遍性并且通过应用STED显微镜的另一项研究证实。另一份报告揭示,虽然树突中也存在肌动蛋白血影蛋白为基础的周期性膜骨架,但树突中这种结构的形成倾向和发育速率低于轴突。此外,本文还显示了胞体和树突部分中肌动蛋白和血影蛋白的2D多边形晶格结构,类似于红细胞中的膜骨架结构。此外,使用SiR-Actin通过STED显微镜可以在活的原代神经元中观察到这种周期性模式。最后,最近的CLEM方法结合铂复制EM研究了无顶轴突中的肌动蛋白组织(PREM)和STORM,并提供了轴突辫状肌动蛋白结构与周期性肌动蛋白超微结构相关的证据(图8)。
图8.CLEM成像结合铂复制电子显微镜和原代神经元无顶轴突的STORM成像通过PREM(灰色)可视化的轴突辫(箭头)映射在大鼠原代神经元的超分辨肌动蛋白环(假彩色)上,比例尺= 2、1、0.2 μ m(从左到右)。中:编织间距的定量显示与周期性肌动蛋白间距相似的尺寸。右图:在PREM记录中评估的微丝厚度,未分裂(编织)和分裂(分裂)的轴突肌动蛋白编织物以蓝色表示,树突中的单个肌动蛋白微丝以紫色表示,微管以灰色表示。给出了平均值和SEM。
β II血影蛋白的敲除导致周期性肌动蛋白环结构的破坏沿着细胞器的双向轴突运输受损。与轴突过程相比,轴突起始节段中基于肌动蛋白血影蛋白的细胞骨架的分子组织与远端轴突相似,并且更稳健,如轴突起始节段(AIS)蛋白锚蛋白-G和β IV-血影蛋白的SMLM所示。此外,β IV血影蛋白和锚蛋白G存在于AIS中,而β II血影蛋白和锚蛋白B存在于远端轴突中。SMLM显示与肌动蛋白环连接的纵向头对头β IV血影蛋白和锚蛋白的二价取向有助于建立致密的AIS超微结构,甚至对靶向肌动蛋白和微管的药物治疗具有抗性。锚蛋白-G进一步显示在神经元活性增加后聚集成亚结构域,并代表精神疾病的主要风险基因。随后的SMLM研究也阐明了α 2血影蛋白与β IV-血影蛋白共同在AIS提供稳健的周期性细胞骨架组织、防止AIS组装不完全以及神经变性的重要性。一份相关报告显示αII血影蛋白丰度与有髓轴突直径成比例,并表明大直径轴突更容易发生神经退行性过程。血影蛋白II免疫染色后,通过ExM研究了β II血影蛋白沿着轴突的周期模式,随后将其连接至可膨胀聚合物并在水中膨胀。这种新方法证实了先前报道的细胞骨架内的组织原则。遗憾的是,在ExM中,鬼笔环肽探针在扩增过程中被洗掉,但是两种策略的途径针对该问题:一方面,携带甲基丙烯酰基的鬼笔环肽的三官能形式被设计用于有效地标记对接到凝胶上;另一方面,最近的一份报告使用了荧光团偶联抗体,该抗体靶向鬼笔环肽探针上的荧光团,该探针可与凝胶连接,类似于常规免疫染色。在中枢神经系统中的几种神经元细胞类型和动物物种中也证实了肌动蛋白和附属蛋白的强大超微结构组织。在周围神经系统中(PNS),STED显微镜也显示梳理的神经纤维制备物上存在重复的细胞骨架成分。最后,SMLM显示了肌动蛋白-血影蛋白骨架的重要生物学功能:它可作为信号平台,通过组织跨膜信号蛋白,包括G蛋白偶联受体(GPCR)、细胞粘附分子(CAM)和受体酪氨酸激酶(RTK),从而实现GPCR和CAM介导的RTK信号传导,实现神经元中的信号转导。
5.2. 突触前室
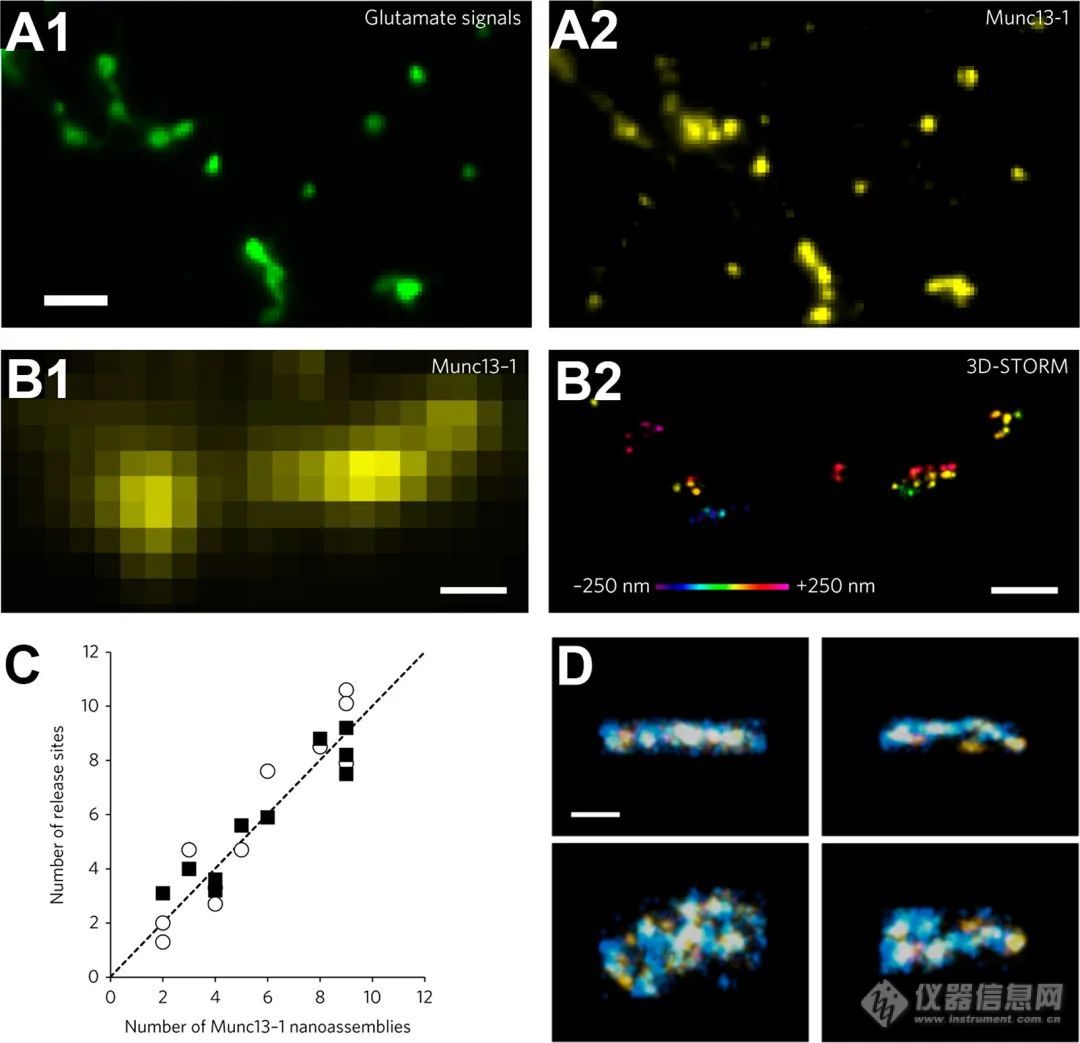
为了确保有效的化学神经传递,突触前结介导突触囊泡循环、神经递质填充以及在称为活性区的专门蛋白质密集纳米区室与突触前膜融合,最终释放其神经递质含量。在此,我们关注SRM如何扩展我们对突触前功能的理解。虽然早期对小直径突触囊泡的研究局限于使用EM化学固定的神经元,随着SRM的出现,突触囊泡在突触前区室中的移动性也可以通过免疫标记钙传感器突触结合蛋白1来可视化(Syt1)驻留在突触囊泡上并应用视频速率STED显微镜。STED显微镜进一步显示,突触囊泡融合后,Syt1分子似乎位于突触膜上,也支持胞吐后突触囊泡蛋白的清除过程。此外,突触囊泡融合期间暴露于细胞外间隙时,荧光团共轭纳米抗体靶向突触小泡蛋白2Hluorin结合后的亚衍射跟踪显示了突触囊泡的异质迁移率。类似方法使用vGlut 1-pHluorin在原代神经元中的表达,以观察单个突触小泡,定位精度为27nm,并显示突触小泡的多个不同释放位点。作为一种方法上的进步,为了可视化活跃的再循环囊泡,设计了一种名为mCLING的亲脂性膜探针,用于染色突触膜,通过内吞作用吸收并固定,可结合免疫标记和SRM。对于突触小泡的胞吐作用,突触小泡在所谓的活性区的膜上的接近和对接以及最终突触小泡的释放需要属于突触前细胞基质的一组突触前蛋白质的高保真度相互作用。黑腹果蝇以其容易获得的基因帮助确定了果蝇幼虫神经肌肉接头(NMJ)活性区的第一个重要蛋白质。Bruchpilot是不可或缺的有源区组件(Brp),是一种大的卷曲螺旋蛋白,对适当的钙通道聚集和突触囊泡定位到突触释放位点至关重要。除了表征Brp的钙通道聚集任务外,STED显微镜还证明了蛋白质的自我组织,并揭示了Brp相互作用蛋白(如syd-1 α、liprin、和rim结合蛋白定量dSTORM方法研究了果蝇活跃区的Brp细丝数量,并显示Brp的结构组织与其功能之间存在很强的相关性。一项后续研究通过Syt 1敲除方法以及随后的电生理学和突触释放研究了活性区组织和突触囊泡释放之间的关系。通过dSTORM评价活动区(CAZ)参数的细胞基质。该研究表明,Syt 1的敲除导致更高的Brp计数和果蝇NMJ 1b型扣簇内Brp图谱的改变。在哺乳动物突触中,突触前支架蛋白巴松管和短笛参与调节突触小泡释放。据报道,巴松管通过与RBP的相互作用来控制CaV2.1型钙通道的定位。此外,巴松管加速小泡释放,因为巴松管的丢失导致突触小泡数量急剧减少和小脑苔藓纤维中的突触抑制至颗粒细胞突触。低音管和短笛的STED显微镜显示三明治结构,短笛在两侧,低音管在中间。脑切片中的STORM成像绘制了低音管相对于脑内其他相关突触蛋白的方向。突触前和突触后区室通过距离测量。囊泡胞吐过程进一步由一组可溶性N-乙基马来酰亚胺敏感因子附着受体(SNARE)蛋白协调。泡状陷阱(vSNARES)和t-SNARES的复合物形成导致突触囊泡融合成功。突触体相关蛋白25(SNAP-25)和突触融合蛋白在质膜上的聚集首先通过STED显微镜进行表征。该研究表明,大约75个突触融合蛋白分子被包装成50至60纳米大小的纳米簇。在后来的研究中,SMLM成像绘制了SNAP-25和突触融合蛋白的分布图,精确度甚至更高。这里,描述了突触融合蛋白簇内分子密度的梯度。dSTORM成像显示,未成簇的分子紧密定位于成簇区域。最近的一项研究产生了靶向突触融合蛋白或SNAP-25的骆驼纳米抗体。这些进入蛋白质密集的突触前区域的效率更高,并显示突触外突触融合蛋白可在增强突触活性后进入突触隔室。SNARE结合蛋白tomosyn被证明位于突触融合蛋白簇,据报道,其β-螺旋桨结构域的突变对与SNAP-25形成复合物至关重要。此外,tomosyn与v-SNARE突触短蛋白竞争与突触融合蛋白和SNAP-25形成复合物,因此被认为下调了胞吐作用。相反,rim结合蛋白2(RBP2)可根据所研究的突触类型在调节突触可塑性方面发挥多种作用。SRM显示RBP2通过调节CaV2.1钙通道相对于释放位点的纳米定位来调节释放概率。此外,STED显微镜显示海马脑片中RBP2对锥体CA3-CA1突触的神经传递只有轻微的影响,它通过控制munc-13-1的定位,在苔藓纤维突触中强烈调节囊泡启动和释放概率。Munc-13是unc-13的哺乳动物直系同源物,是启动突触小泡释放的另一个不可或缺的组分。Munc-13-3被证明可将钙通道募集到活性区,3D-STORM成像提供了munc-13-1在突触释放位点聚集并与突触融合蛋白分子结合的证据(图9),通过实时成像和STED显微镜研究果蝇中与突触前钙通道距离不同的两种munc-13亚型,揭示了unc-13亚型通过CAST定位。ELK同源物Brp和RIM结合蛋白,导致释放位点相对于钙通道的不同超微结构布局。Unc-13异构体在神经元发育过程中改变其比例,通过改变其与钙通道的纳米结构域偶联而导致神经传递增强。通过突变分析进一步分析了Unc-13 A的定位,提供了C端部分调节释放位点生成而蛋白质N端部分参与活性区靶向的证据。
图9.与突触释放位点相关的Munc-13 - 1纳米组装体
上排:通过应用高钙溶液和钾通道阻滞剂(A1)和相应的munc-13 - 1信号(A2)进行突触激活后eEOS(增强的谷氨酸光学传感器)的荧光响应的图像。注意在主动释放条件下的紧密空间相关性。比例尺:5 μ m。第二行:munc-13 - 1的常规显微镜图像(B1)与3D STORM成像(B2)。比例尺:500 nm。(C)释放位点数量与munc-13 - 1纳米簇数量的相关性。(D)munc-13 - 1(黄色)和突触融合蛋白-1A(蓝色)的超分辨双色可视化。比例尺:2 μ m。侧视图(顶视图)和正面(底视图)。比例尺:200 nm。
在活动区内,钙通道的纳米定位和潜在的动力学对于神经传递的有效调节至关重要。钙通道和突触囊泡之间的纳米结构域耦合可以紧密或松散,并以这种方式形成神经传递保真度。小脑突触中的免疫金EM定量显示,强突触显示耦合距离约为10 nm,但与较弱突触相比,其钙通道少三倍,较弱突触的特征是效率较低。更长的偶联距离。免疫金冷冻断裂EM研究结合电生理学提供了释放位点与突触前钙通道数量成比例的证据。为了表征电压门控钙通道的迁移率,在原代神经元中应用单粒子示踪PALM(sptPALM)成像。研究表明,集中在突触前位点的通道中有60%是移动的。此外,通过应用BAPTA进行钙缓冲降低了钙通道的扩散。结果表明,突触囊泡和钙通道之间的非随机域偶联形成了神经传递保真度,并可根据需要通过突触前钙通道的扩散进行精细调节。融合和递质释放后,内吞机制诱导循环产生新的囊泡,从而重建可释放囊泡库并为持续神经传递提供基础。囊泡循环的主要机制由网格蛋白介导的内吞作用组成。使用光遗传学和闪光-冷冻电子显微镜的研究也报告了超快内吞过程快200倍。通过摄取Syt 1抗体(靶向囊泡上的管腔结合位点),将内吞位点映射到围活动区,如双色iso-STED显微镜所示。此外,在神经内分泌细胞中,STED显微术还揭示了囊泡仅能部分地与突触前膜融合以通过形成Ω形结构而不完全并入膜中来释放递质,因此有利于递质释放的"吻和跑"模型,与网格蛋白介导的内吞作用相比,该模型导致快得多的囊泡再循环速率。依赖于活动的大量内吞作用进一步增加了可能机制的复杂性,有人认为多种内吞作用模式可能会根据突触类型和活动平行运作。
5.3. 突触后室
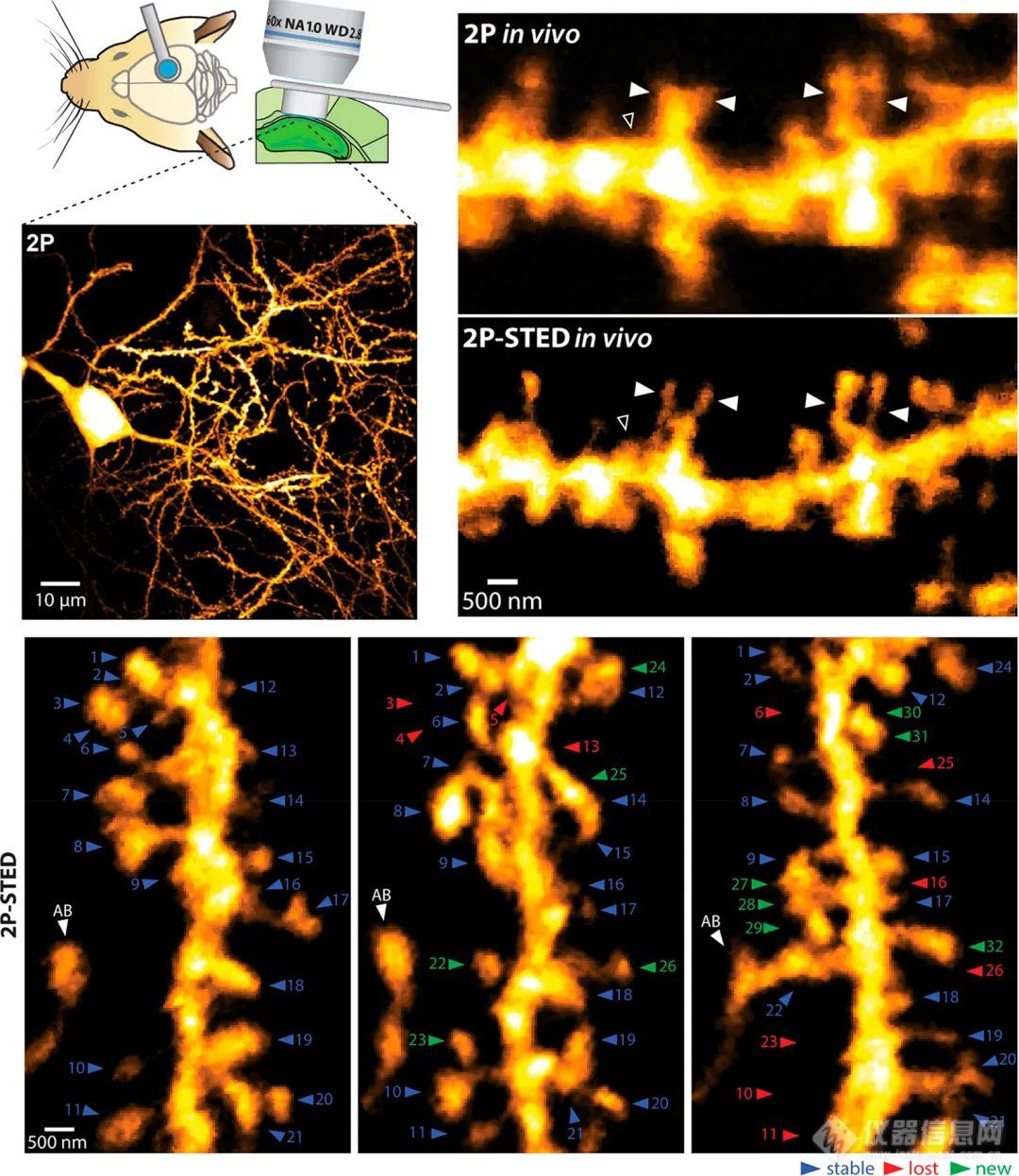
突触后区室作为突触受体场,负责传递来自突触前终末的信号。它结合了负责锚定突触后受体的支架蛋白和专门用于信号整合的信号分子。在神经元树突中,由小体积树突棘提供区室化,树突棘起源于主树突轴并从主树突轴突出,并根据突触活动和发育阶段显示大小和形状的动态变化。树突棘作为突触长时程增强(LTP)的结构相关物,因此与学习和记忆有关。其体积小、形状多面、动态特性要求采用基于光学显微镜的方法,分辨率超过衍射极限,并具有实时成像的潜力。在原代神经元中应用livecell SMLM的首批研究之一使用碳菁染料如Dil使棘和丝足可视化。为了使突触后膜结构可视化,已经提出了新的膜标记剂工具箱,其使得能够对树突棘进行神经元追踪和可视化。最近,已经通过快速SIM和增强共焦显微成像研究了称为小刺的树突棘上的微小突起的动力学。通过结合SIM成像和计算方法进一步评估树突棘的几何形状,以表明凹面对棘结构稳定的重要性。在树突棘中,肌动蛋白高度富集,并显示主要定位在突触后密度(PSD)和棘膜。肌动蛋白分子速度的升高已被绘制到整个棘(棘尖除外)的亚结构域。为了分析脊柱中肌动蛋白的动力学,设计了一种低亲和力光转换肌动蛋白探针,并通过STED显微镜使用减少畸变的光学器件表征了活脑切片中的动力学。通过应用鬼笔素ATTO 647 N标记的原代神经元的STED显微术,在树突棘颈以及丝状伪足样棘中观察到个周期性的F-肌动蛋白片段。同年,STORM成像还显示树突棘颈和丝状伪足样棘中存在基于肌动蛋白的周期性膜骨架。在树突棘中,分支的F-肌动蛋白在PSD附近成核,而伸长仅限于指状突起的尖端,并为棘突提供基础。树突区室中肌动蛋白组织的异质性也可以通过应用基于监督学习的模式识别分割来自动分析。这种方法已经在树突状F-肌动蛋白的SMLM数据上进行了测试,并使用树突的铂复制EM进行了验证。活体小鼠脑切片中的延时STED显微镜结合FRAP和海马CA1神经元的电生理学可以提供证据,证明脊柱颈部是可塑性元件,在神经递质释放诱导的长时程增强后,其宽度增加,长度减少。使用直立STED显微镜获得了活体小鼠脊柱动力学的第一个SRM。在此,Thy1-EYFP小鼠躯体感觉皮质中的脊柱在其头部和颈部表现出形态可塑性。还使用慢性2P-STED成像研究了活体小鼠的海马脊柱动力学,显示脊柱密度比早期报告高2倍,并提供了几天内脊柱更新率的估计值(图10)。
图10.海马CA1锥体神经元脊柱转换的体内长期2光子STED成像
左上图:使用长工作距离物镜和CA1锥体神经元的双光子概览图像的实验方法。右上角:常规2-光子成像与2-光子STED成像的比较揭示了总体上更高的棘密度和更详细的形态,尤其是在轴和棘突起中。空心箭头表示传统双光子成像无法分辨的棘突,而实心箭头表示双光子STED报告棘突数量更多且形态更复杂的棘突。底部图像:在CA1海马区基底树突的选定区域中的脊柱更新成像几天(说明了第0、2和4天)。脊椎连续编号。AB =接近树突的轴突结(回缩的棘标为红色;绿色的新刺)。
此外,sptPALM揭示了突触后激酶CaMKII的空间和动力学亚群,CaMKII在突触棘中富集并介导Ca 2+依赖性可塑性机制。这些动力学似乎是由棘肌动蛋白调节的,因为Latrunculin A导致了棘内CaMKII扩散的显著变化。在PSD内的棘头处,一个密集的蛋白复合物包含了不同的突触后支架蛋白,如PSD-95、homer 1和shank 3,这些蛋白排列在突触下结构域中,大小约为80 nm。PSD-95被证明是动态组织成单个单位或由多个纳米簇组成,这取决于突触类型。突触后支架蛋白的STED显微镜揭示了它们负责将离子型受体锚定到突触后膜上,如活的初级神经元的SMLM所示。在此,活细胞单分子成像结合定量分析揭示了含谷氨酸A2的AMPA-R优先聚集在突触下PSD-95簇的稳态调节。AMPA-Rs的侧向扩散也与突触的短时程增强和长时程增强(分别为STP和LTP)相关。例如,研究表明,为了从突触抑制中恢复,脱敏受体通过横向扩散与功能受体交换。此外,跟踪实验表明,在CaMKII激活诱导LTP后,AMPA-R扩散捕获到突触部位。该过程由钙浓度升高触发,导致CaMKII介导的司他加嗪磷酸化,其与PSD-95一起能够调节AMPA-R迁移率。进一步的研究报道AMPA-Rs的交联导致受体在膜中的不动性并阻止成功的LTP诱导,这一机制也可能与AMPAR致病性抗体介导的自身免疫性中枢神经系统疾病的病理生理学有关。AMPA-R的纳米级组织以可变簇大小为特征,也与NMDA-R的GluN1亚基和mGluR 5代谢型受体进行了比较。令人惊讶的是,突触前mGluR5受体表现出更均匀的分布,没有聚集行为。代表NMDA-R总库的含内源性GluN 1亚基的受体显示聚集在一个主要的单簇中,该簇被单独的受体包围,这与新型基于敲入的基因组编辑方法的观察结果相似。当关注NMDA-R的不同亚型(细分为含NMDA-R的NR 2A和NR 2B)时,SMLM显示,在突触发育过程中,它们被分割成纳米结构域,并根据其突触比例进行重塑。关于谷氨酸受体的迁移率,单分子追踪实验揭示,神经元活性优先影响AMPA-R的迁移率,而NMDA-R的移动是由蛋白激酶C活性触发的,而不是由钾升高触发的。此外,dSTORM成像表明,不同的NR 2亚单位定位于不同的纳米结构域,这些纳米结构域在神经元发育过程中表现出灵活性。根据NR 2A和NR 2B的纳米结构,LTP表达可以双向方式调节。红藻氨酸盐受体的单分子追踪实验也显示了突触活性增加后的突触捕获。此处,突触激活诱导红藻氨酸盐受体与突触β-连环蛋白/N-钙粘蛋白复合物结合,形成短期可塑性。
作为抑制性突触的对应物,gephyrin是将GABAA(GABA-A R)或甘氨酸受体(GlyR)整合到突触后膜所必需的关键锚定分子。通过PALM/dSTORM成像对脊髓突触的gephyrin进行分子计数,结果显示抑制性PSD(iPSD)体积为0.01至0.1 μm3,每个iPSD有200− 250个gephyrin分子。单分子成像进一步揭示了gephyrin分子与受体结合位点的化学计量比约为1:1.96。类似于兴奋性突触,抑制性PSD(iPSD)根据突触活动动态调节其大小。通过NMDA-R激活的抑制性突触的LTP导致突触gephyrin积累增强,允许以CaMKII依赖性方式增加GABA-A R聚集,从而诱导GABA能突触后电流增强。相比之下,抑制gephyrin募集到突触区域导致GABA-A R迁移率降低,并阻止iLTP诱导。iLTP诱导后,gephyrin分裂为纳米结构域。gephyrin的重组降低了抑制性突触后电流的振幅变异性,因此为正确的GABA-A R定位对iLTP真正表达的重要性提供了证据。有趣的是,单粒子追踪显示脱敏的GABA-A R甚至可以通过横向扩散在并列的GABA能突触之间交换,提供了控制GABA能电流的另一种机制。此外,为了阐明多巴胺能突触的超微结构布局,dSTORM成像将多巴胺转运蛋白映射到胆固醇依赖性纳米结构域,为更好地理解多巴胺能神经传递的病理生理过程奠定了基础。
5.4.亚突触结构域中的跨突触排列
突触强度取决于突触前融合位点和突触后受体组织之间的空间关系的观察结果已经在早期的电生理学实验中提出,这些实验描述了释放取决于释放位点的数量、突触囊泡的释放概率以及受体提供的突触后基本反应。这种亚结构域的跨突触组织首先被观察到标记边缘的突触粘附分子SynCAM1和位于PSD更中心的EphB2。SynCam1在PSD中形成突触下云,可通过长期抑郁模式重塑。29SMLM结合链霉亲和素的新单体变体(旨在减少突触区域的交联和空间位阻),用于显示跨突触伙伴神经木质素1和神经肽1ß在突触处扩散性捕获,形成相对簇。近年来,兴奋性突触的活性区细胞基质和突触后受体-支架集合的跨突触排列被发现与靶向神经递质释放有关,从而提供高保真的突触传递。在这里,释放位点定位是通过一种新的方法实现的,该方法基于融合到突触小泡蛋白Vglut 1的pHluorin标签和RIM 1/2纳米簇的超分辨检测。多色三维定位显微镜显示RIM 1/2和突触后PSD-95形成相对的纳米团簇。LTP诱导导致PSD-95密度碎片增加,并增强了此类纳米柱的排列,而LTD方案导致突触后柱的解体。突触前和突触后关键分子的纳米级排列归因于神经木质素1。此外,在应用STED显微镜的实时成像实验中报告了棘体积增加与排列纳米模块数量之间的紧密相关性。也报告了抑制性突触的突触下结构域纳米级排列。STED和SIM揭示了金卟啉与GABA-AR突触下结构域的紧密联系。此外,突触后GABA-A R显示与突触前RIM结构域共排列(图11)。在鼠类神经肌肉接头处,通过应用SIM成像,突触后乙酰胆碱受体与接头褶皱和突触前活性区开口的排列已经可视化。
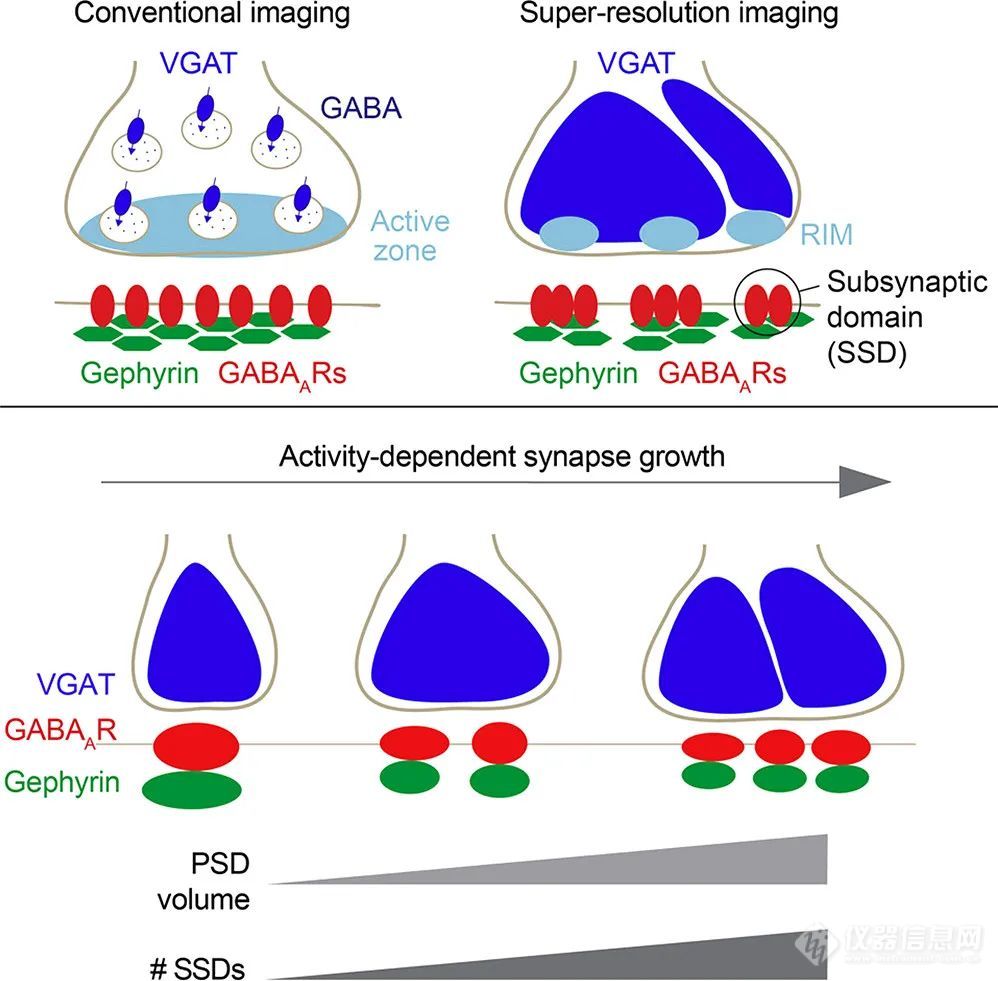
突触前RIM元件与突触后金卟啉支架分子和抑制性突触处GABA-A R突触下结构域的排列PSD的体积和突触下结构域的数量与活动相关的突触大小变化成比例。
5.5. 三联突触
星形胶质细胞是神经传递的基本调节者,神经元突触周围突触前星形细胞突起(PAPs)的吞噬产生了三联突触这一术语。PAP能够通过传递调节分子来修改和控制突触传递。通过dSTORM重建星形胶质细胞过程,可通过标记胶质酸性纤维蛋白来实现星形胶质细胞的纳米可视化(GFAP)和谷氨酰胺合成酶和S100 b的成像。最近的一份报告应用ExM在脑切片中显示突触周围星形胶质细胞谷氨酸转运蛋白,显示在较大的神经棘中谷氨酸摄取效率降低,这与这些棘的较高GLT-1水平相关(图12)。
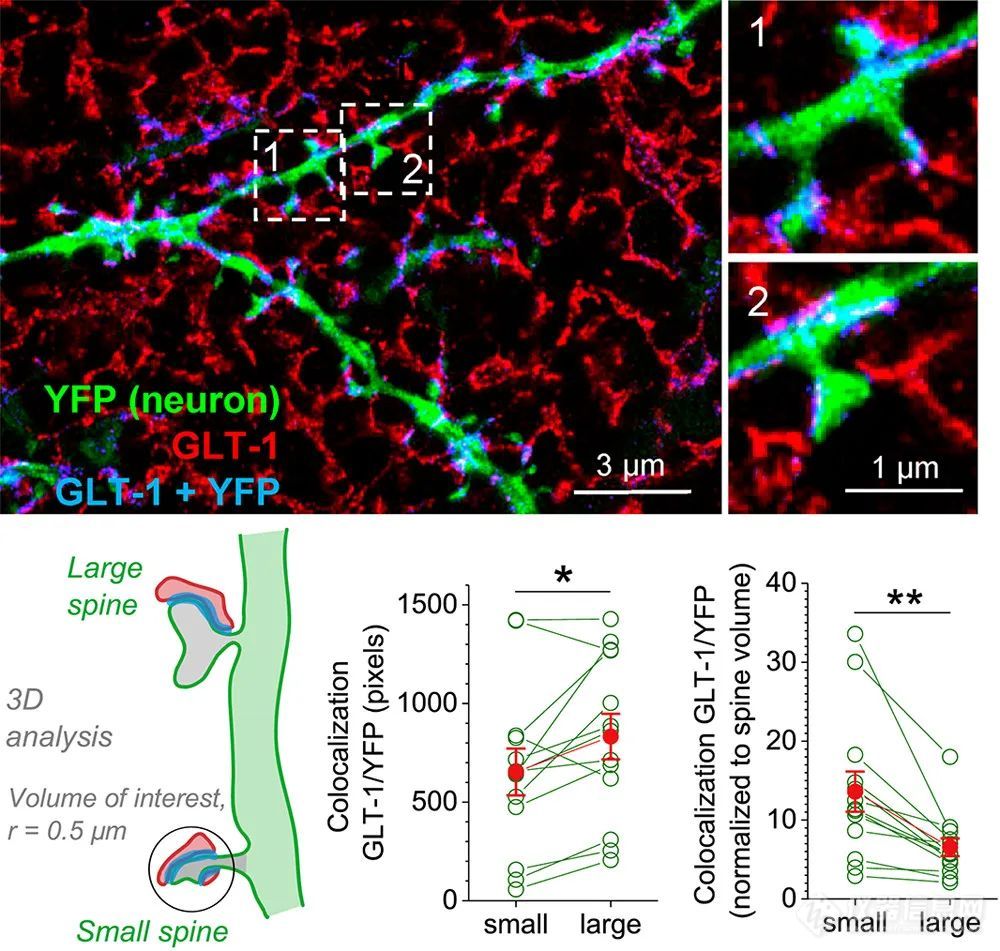
图12.海马脑片中CA 1锥体神经元周围星形胶质细胞突起的ExM使用Thy1-YFP小鼠系标记锥体神经元树突(绿色);星形胶质细胞通过GLT-1免疫染色(红色)显现。蓝色信号代表树突区室和星形胶质细胞过程的共定位。右侧面板中提供了更高放大倍数的插图。左下角:大小刺的分类原则。底部中间和右侧:GLT1和神经元YFP的共定位像素的定量。注意右侧脊柱体积归一化后的变化;红点表示平均值和SEM,p = 0.0220(绝对GLT-1覆盖率),p = 0.00223(相对GLT-1覆盖率)。
EM和STORM发现,PAP还配备了局部翻译位点,以避免星形胶质细胞体细胞中合成的蛋白质的长运输路线。最近对器官型切片进行的3DSTED显微镜研究揭示了星形胶质细胞钙信号的结构先决条件。在星形胶质细胞内检测到海绵状结构域,其包含与突触位点非常接近的节点和轴。钙瞬变的共聚焦成像与星形胶质细胞结构的STED显微镜检查的相关性显示,自发钙瞬变紧密地映射到这些节点。因此,这些结节被认为是类似于树突棘的空间分隔。胶质递质的胞吐作用需要提供胶质囊泡。通过将电容测量与摄取葡聚糖后星形胶质细胞内囊泡的SIM图像相关联,揭示了外吞和内吞之间界面处的动力蛋白依赖性膜中间体。通过STED显微镜和SIM分析单个神经胶质囊泡,表征了星形胶质细胞中大小和融合能力不同的两个囊泡群。根皮素实验结合SIM测定星形胶质细胞囊泡上Syb2分子的拷贝数为25。此外,使用STED和TIRF显微镜在单个囊泡水平分析培养星形胶质细胞中的VAMP3阳性囊泡。测量结果显示,VAMP3覆盖的囊泡大小约为80 nm,并提供了这些囊泡参与钙依赖性囊泡循环的证据。SIM成像还可揭示突触泡蛋白(一种已知参与神经元胞吐作用的v-SNARE蛋白)也普遍存在,并组织在单个星形细胞囊泡上以进行有效胞吐作用。星形胶质细胞也参与促进兴奋性LTP通过再循环proBDNF到BDNF。这里,SIM成像显示proBDNF位于体细胞区域的囊泡大小的簇中,而点状模式主要沿着星形细胞端足,以扩大BDNF在记忆形成中的作用。为了尽量减少激发光的散射,采用被动CLARITY方案和多光子显微镜,通过组织清除改善组织深处星形胶质细胞的成像。通过STED显微镜(使用SiR-肌动蛋白和SiR-微管蛋白探针)和原子力显微镜(AFM)的相关方法,将星形胶质细胞骨架和膜的生物物理特性联系起来,以测量膜拓扑结构和刚度。

目前在国内,随机光学重建显微镜STORM已成功实现商用,有需要STORM成像技术进行实验研究的专家老师们,可预约获得 iSTORM 超高分辨率显微成像系统试拍服务~
 图、iSTORM拍摄的超高分辨率细胞图片
图、iSTORM拍摄的超高分辨率细胞图片
力显智能现已发布的超高分辨率显微成像系统 iSTORM,成功实现了光学显微镜对衍射极限的突破,使得在20纳米的分辨率尺度上从事生物大分子的单分子定位与计数、亚细胞及大分子复合物结构解析、生物大分子生物动力学等的研究成为现实,从而给生命科学、医学等领域带来重大突破。
About us
宁波力显智能科技有限公司(INVIEW)是专业从事超高分辨率显微技术和产品研发的科技企业,依托复旦大学的自动控制、新一代信息技术及香港科技大学的生物、光学、图像处理等的技术,拥有光学、生物、自控、机械、信息技术等多领域交叉学科技术团队,将2014年诺贝尔化学奖技术产业化,推出了超高分辨率显微成像系统iSTORM、细胞智能监控助手赛乐微等一系列产品,帮助人们以前所未有的视角观察微观世界,突破极限,见所未见。

 关注
关注