
 关注
关注
已关注
![]() 已认证
已认证
粉丝量 0
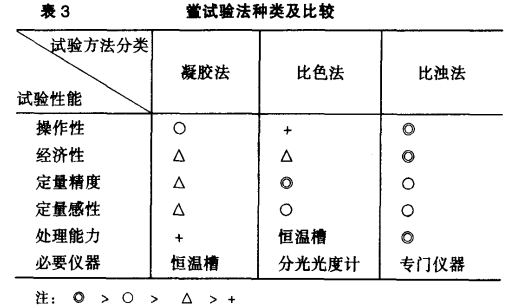
鲎试验法种类及比较
根据反应机理的不同又可将鲎试验法分为以凝胶的形成作为指标的凝胶法,以凝胶的浊度变化作为指标的比浊法以及显色合成基质水解显色作为指标的比色法,其中以定性的凝胶法收载最为普通(表3)。 根据反应机理的不同,主要可分为以下几类:(1)凝胶法(Gel-Clot technique):可在0.03Eu/ml的范围内进行半定量测定。(2)比浊法(Turbidimetric Assay):通常在0.006 ~ 300Eu/ml的范围内进行定量。(3)比色法(Colorimetric Assay):操作比较复杂,但灵敏度和精确度高。通常在0. 006~15Eu/ml的范围内进行定量。(4)ELISA(Enzyme-Linked immunosorbent assay):用鲎试剂与酶联吸附免疫测定法联合检定微量内毒素的方法。(5)以巨噬细胞产生的细胞分裂素(Cytodines):作为机体反应指标的热原检查系统。 凝胶法:即在100 x 75mm的小试管内,加入待检样品的鲎试剂混合。37℃60分钟保温反应后,180°倒转,根据凝胶形成的有无作为判定指标的一种半定量法。此法操作比较简单、经济,而且不需要专用测定仪器。可在0.03Eu/ml的范围内进行半定量测量。 比浊法:根据内毒素与鲎试剂的(C因子)反应机理,被激活的凝固酶切断凝固蛋白原中特定位置的精氨肽链。形成凝固蛋白,产生凝胶的浊度变化,适当的浊度测定仪进行定量的一种方法。可分为Endopoint Assay和Kinetic turbidimetric Assay。此法操作简单、经济、灵敏度高、检测范围广。通常在0. 006~300ET/ml的范围同时进行定量。但需要专用仪器。 比色法:根据被内毒素激活的凝固酶水解鲎三肽中精氨酸肽链。释放出对硝基苯胺(PNA黄色A405nm)用冰醋酸中止反应进行吸光度测定,或用游离的PNA和偶氮试剂反应,最终形成偶氮蓝复合物显色(红色A 545nm)而进行比色测定的一种方法。可分为Endopoint Assay和Kinetic turbidimetricAssay。此法操作比较复杂。但灵敏度、定量性和精度都高。通常在0. 006 ~15Eu/ml的范围内进行定量,而且更微量的内毒素也可定量。目前主要用在血液﹑体液中的微量内毒素的测定方面。
参数原理
2022.11.15
内毒素效应中起主要作用的炎症因子
内毒素是革兰氏阴性杆菌细胞壁的主要成分,对机体代谢的影响如对糖的利用障碍、氨基酸的消耗增加都是通过细胞因子实现的。在内毒素介导产生的诸多激素和介质中,最重要的是来自免疫系统和血管内皮细胞系统的细胞因子,主要包括肿瘤坏死因子、白细胞介素-1、白细胞介素-6、白细胞介素-8等。一般认为,单核巨噬细胞是产生细胞因子的主要细胞,故有“细胞因子工广”之称。 内毒素在创伤后机体的各种反应中起了触发剂的作用,在激活产生的细胞因子中,肿瘤坏死因子-α可能起核心作用。其证据是注入肿瘤坏死因子后机体几乎可出现脓毒症的所有症状,用抗肿瘤坏死因子的抗体可以保护机体免受致死量脂多糖的损害。肿瘤坏死因子和脂多糖生物学特性有许多相似之处。机体在对脂多糖的反应中,肿瘤坏死因子-α是早期出现的细胞因子。肿瘤坏死因子-α在炎症反应中是激活细胞级联效应的主要介质,当细菌或脂多糖侵入机体后,肿瘤坏死因子-α在循环中可出现较早并迅速达到高峰,尽管半衰期很短,但足以诱导“次级”细胞因子(如白细胞介素-1、白细胞介素-6等)产生。 白细胞介素-1使中性粒细胞聚集和发生急性反应,它可由内毒素直接刺激巨噬细胞,也可由肿瘤坏死因子作用于巨噬细胞或通过刺激内皮细胞产生。白细胞介素- 1可引起T细胞增殖,诱导可溶性白细胞介素2受体的产生及增加可溶性白细胞介素受体的表达,和白细胞介素-4一起激活细胞,使其增殖和分泌免疫球蛋白。 肿瘤坏死因子-α和白细胞介素-1间有相互调控、促进或诱发释放作用。动物实验表明肿瘤坏死因子有诱导白细胞介素-1、白细胞介素-6合成的作用。另外,白细胞介素-1也可以诱导白细胞介素-6合成。白细胞介素-6还可由严重感染或脂多糖及其他一些细胞因子如白细胞介素-3、干扰素、前列腺素-F和集落刺激因子等诱导产生。在各种类型的腹部手术病人中发现血清白细胞介素–6水平和代表炎症反应的C-反应蛋白(CRP)关系密切。白细胞介素-6和白细胞介素-1有协同作用,可促进未被刺激的B细胞生长和分化。在创伤或内毒素血症的病理过程中,白细胞介素-6可大量产生,其水平与疾病的预后及患者的死亡有明显关系。 白细胞介素-8是另一种参与炎症反应的细胞因子,其浓度高低与疾病的严重程度相关。白细胞介素-8是一种低分子量的细胞因子,有激活中性粒细胞的作用。Kimberly等证明在注射脂多糖或白细胞介素-1后,当白细胞介素-8浓度达最高时,循环中中性粒细胞数从最初的粒细胞减少很快恢复,可能是由于白细胞介素-8能够减少中性粒细胞粘附到内皮细胞上;它还可在炎症过程中引起中性粒细胞向炎症部位集聚。另外,它还能调节血管内皮的通透性。白细胞介素-8既可由脂多糖刺激产生,亦可由肿瘤坏死因子和白细胞介素-1刺激产生。早期局部产生的肿瘤坏死因子和白细胞介素-1可能有助于白细胞介素-8的表达。 白细胞介素-8还可减少急性炎症时多核巨细胞的聚集,所以白细胞介素-8可能有减轻炎症的作用。在炎症反应发生后,体内出现作用广泛而复杂的细胞因子,它们之间形成了细胞因子网络,这个网络具有调节作用,其中有的起促进(上调)作用,另一些则起抑制(下调)作用,从而而使这一系统更加完善。
参数原理
2022.11.14
鲎试验法的反应机理简介
鲎试验法也称为细菌内毒素检查法,英文Bacterial Endotoxin test,系列用革兰氏阴性菌所产生的内毒素与TAL或LAL试剂发生反应,根据所形成的凝胶化反应,从而定性或定量检测细菌内毒素的一种体外分析方法。有关细菌内毒素与LAL试剂或TAL试剂的反应机理研究,虽然在研究鲎血凝集反应的初期,leven and Bang就已经初步阐明了凝集反应的机理;但进一步从分子水平上的研究工作,一直到70年代后期才逐渐被阐明。鲎试验凝集反应的机理如下图所示: 即鲎血变形细胞裂解物中含有能被内毒素激活的凝固系统,主要有凝固酶原,C因子、B因子与凝固蛋白原等,内毒素激活C因子,活化的C因子又激活B因子(C因子系),或由1,3)-β-D-葡聚糖激活G因子(G因子系)接着激活的B因子或激活的G因子,再去激活凝固酶原,使其转变成活化的凝固酶,该酶切断凝固蛋白原中特定位置的精氨肽链,形成凝固蛋白产生凝胶。关于G因子系能否单独成立,目前在各国存在分歧,凝固酶除能水解凝固蛋白原使之变成凝固蛋白而发生凝固外,它还对外加的鲎三肽有去胺酶活性使硝基苯胺(pNA)游离,而发生呈色反应。
参数原理
2022.11.14

《中国药典》药物制剂质量控制技术研讨培训完美收官!
11月11日,在《中国药典》药物制剂质量控制技术研讨线上培训最后一天,科德角国际生物医学科技(北京)有限公司资深技术总监范玉明受邀参会并做研讨培训发言。 此次线上研讨培训主要为进一步强化药品生产全过程管理理念,帮助行业掌握《中国药典》相关技术要求,加强对药品生产用原辅包以及生产过程的控制,进一步保障药品质量,邀请药典委员会相关专业委员会委员、药检机构、医药企业等医药领域具有广泛影响力的代表进行研讨交流,共同对《中国药典》新型药物研制质量控制考虑要点等相关技术要求进行全面深入解读;探讨先进检测技术在药品质控的应用以及分享新型药品生产工艺风险管控策略的经验。 科德角国际技术总监范玉明在研讨培训中,聚焦细菌内毒素检测技术领域,介绍了热原检测与其替代方法发展史、细菌内毒素光度测定法优化升级后的优势以及天然LAL与rCR比较等,还对科德角国际细菌内毒素检测实验室质量控制作了着重阐述,分享了科德角国际细菌内毒素检测实验室的检测实例,为广大线上听众带来了一场内容丰富、干货满满的研讨培训内容。 科德角国际自成立以来,一直专注于细菌内毒素检测领域的全方位服务,始终为客户提供优质的产品及专业的技术服务,着力推动细菌内毒素检测领域高质量发展。 扫描上方二维码观看回放内容↑
参数原理
2022.11.14

内毒素的致病机理简介
业内已证明,细菌及其毒素可刺激体内单核巨噬细胞系统合成、分泌大量细胞因子,并通过网络作用,最终导致过度的全身炎症反应及组织器官损害。研究表明,内毒素是介导革兰氏阴性菌脓毒症的重要启动因子,通过其结合受体或调节蛋白的作用诱导宿主多种细胞因子的合成和释放,激发机体一系列病理生理改变(图13)。 内毒素通过一些中间介质发挥毒性作用。目前多数学者认为单核巨噬细胞在内毒素发挥毒性作用中起重要的介导作用。即内毒素通过与单核巨噬细胞膜或多形核白细胞上的特异性受体CD14。相结合,刺激单核巨噬细胞或多形核白细胞产生并释放一级介质,如肿瘤坏死因子、白细胞介素-1、白细胞介素-6、白细胞介素-8等,从而产生全身急性炎性反应。上述一级介质在刺激机体产生炎症的同时,也可引发二级介质,如前列腺素、血小板活化因子、一氧化氮、弹力酶等的释放,加重原有的病理反应,共同诱发炎性反应综合征、脓毒综合征、内毒素休克、多器官功能衰竭等临床常见急危重性疾病。
参数原理
2022.11.11

影响宿主对内毒素反应的因素
由于人类种族、性别、地域、所处地理环境等不同,使得人们抵抗外界刺激侵袭的能力也不相同,因此对内毒素的反应也存在着个体差异。以下几种因素影响宿主对内毒素的反应。 1、杀菌性/穿透性增强蛋白(BPI),来自中性粒细胞,通过与血清脂多糖结合蛋白(LBP)竞争性结合脂多糖,抵消内毒素在循环中的毒性作用。 2、抗脂多糖抗体(IgM)结合并灭活那些宿主已接触过的异性脂多糖。 3、高密度脂蛋白(HDL)代谢并清除脂多糖。 4、脂多糖结合蛋白运送脂多糖到炎症部位,在此启动炎症细胞因子反应,在任何部位中,脂多糖结合蛋白的数量决定了反应的程度。例如,在一个脓肿部位,中性粒细胞数量占优势,此部位的杀菌性/穿透性增强蛋白的比例高于脂多糖结合蛋白,因此在这一部位释放的脂多糖会被中和。相反,如果脂多糖释放入关节、脑脊液或腹膜液,则更可能与脂多糖结合蛋白而不是和杀菌性/穿透性增强蛋白相互作用,因此激活炎症细胞因子系统。 5、宿主肿瘤坏死因子(TNF)的产生依赖于遗传因素,调节肿瘤坏死因子产生的等位基因纯合子会产生高水平的肿瘤坏死因子。所以在这些个体中,败血症往往是更加致命的。
参数原理
2022.11.11
关于鲎试验法中假阳性结果的解决方法
样品中混入细菌内毒素发生凝集,由形成凝胶的有无来检测样品中的内毒素,然而鲎试剂除了内毒素反应外,它同时也会对内毒素以外的其他物质存在凝集反应。因此,势必会形成假阳性或假阴性的实验结果。这就是鲎试验法中的特异性问题。 对于假阳性结果的问题,可以说十分复杂。因为对于大多数样品来说,主要都为假阴性情况较多,而对于假阳性结果的判断和排除是很困难的。当我们对一个样品用鲎试验法检测出阳性结果时,一可以说是由于内毒素超过限值所引起,二也可以理解为假阳性结果。因此我们在实验时要特别注意区分。 在假阳性结果的研究过程中,最突出的是有关G因子反应系被激活所形成的凝胶化反应,即一般的鲎试剂由于含有G因子也可被(1,3)-β-D-glucan(葡聚糖)所激活而使产生凝集反应,从而形成假阳性结果。关于(1,3)-β-D-glucan:它是在1981年由Adinuma所发现一种抗癌药,能使鲎试剂产生凝固。国外报道该物质在10ng/ml时就可使鲎试剂凝固。根据我们的试验发现不同厂家的鲎试剂可在2.5 ~10ng/ml范围内使鲎试剂凝固。该物质广泛存在于担子菌、真菌、地衣类、酵母和藻类的菌体成分中。另外,某些用人造纤维造的人工肾透析膜上也含有类似物质,但它可引起家兔升温。 (1,3)-β-D-glucan对热极为稳定。高压121℃并不能使其灭活。需250℃2小时加热灭菌才能使其灭活。因此,要想防止假阳性结果的产生,就必须要排除(1,3)-β-D-glucan,使其不能激活G因子而形成凝胶。国外在80年代后对此进行了许多的研究和改进。目前已开发出仅对内毒素起反应的专一鲎试剂。只与内毒素起反应的鲎试剂有:日本生化学工业株式会社生产的Endospecy,它是将东方鲎裂解液中各组分提取后重组,重组时不加G因子,另一种是日本和光纯药株式会社生产的ES =Ⅱ,它是美洲鲎裂解液中加入大刃羧甲基curdaln封闭G因子。同时国外也开发出只与(1,3)-β-D-glucan起反应的鲎试剂:由蚕中提取而来的新型的SLP试剂。(和光)以及日本化学工业株式会社生产的Gluspecy。 对于大多数样品来说,由于使用特异性鲎试剂与非特异性鲎试剂对其结果是没有差异的。因此在一般情况下可以不必考虑,只有在确实认定有干扰时,才考虑使用特异性鲎试剂以排除干扰。
参数原理
2022.11.11

鲎试验法中的干扰因素有哪些?
由鲎试验的反应机理我们可以知道:内毒素在二价阳离子参与下,作用凝集酶系统而发生一系列生化反应,最后形成凝胶。因此,一切阻碍反应机制的因素,均干扰凝胶的形成,形成假阴性结果,作为干扰凝集反应的物质主要有:人血中的多种凝血因子、很多中药成分、某些生化药品、抗生素、可使蛋白质变性的物质、高糖溶液、络合物等等,此外样品的pH值,反应温度等也都有一定的影响(表2)。 另外一些金属离子(如 Fe、Al、Ca)会对细菌内毒素活性产生影响,从而出现假阴性结果,因此在实验时,避免使用一些金属用具,而使用玻璃用具。 关于供试品中自身成分的影响,可以说是鲎试验中影响因素最大的,作为供试品,既有对LAL或TAL试剂的活性产生影响的因素,也有对细菌内毒素的活性产生影响因素,因此在实际测定时要引起注意。 (1)供试品对鲎试剂的活性产生影响的因素为:蛋白酶、蛋白酶抑制作用,螯合剂、蛋白变性剂等。 (2)供试品对细菌内毒素的活性产生影响的因素:金属离子(铁、铝、锗等),表面活性剂等。 (3)对主反应以外的测定系统产生影响的场合:浊度、颜色、磺胺制剂(合成基质化的偶氮化反应时)等。 在这些共存物质的影响当中,特别要引起注意的是些外界金属离子的影响,尤其是在旧菌内毒素标准品的溶解及稀释过程中一定特别注意金属离子的混入,最好的方法是使用玻璃制备的盖子。 对于大多数的注射剂一般在原液时均存在干扰作用,然而当把样品用水进行稀释后即可排除干扰,而对于稀释法不能排除干扰的样品,可考虑用其他方法给予解决。如:中和法、过滤法以及透析方法等,目前市场上见到鲎试剂主要为TAL试剂和LAL试剂。因为工艺不同,抗干扰能力的强弱不一,因此对同一样品、使用不同厂家的LAL试剂所出现的最大无干扰浓度是不同的,这一点要引起注意。另外,TAL试剂和LAL试剂是否完全等活性关系还有待进一步研究和探讨。
参数原理
2022.11.10

鲎试验法的出现和发展是怎么样?
众所周知,美国是发现、发明鲎试剂和最先建立鲎试验方法的国家,早在1956年,美国动物学家于F·Bang 发表了一篇《鲎的一种细菌性疾病》的论文,详细地阐述了细菌毒素使蕴血凝固的现象,随后F·Bang和J·Levin合作开始了对鲎凝血研究,并在1964年提出了鲎血凝集的初步机理,到了1968年两人终于建立了鲎试验方法。 其后随着人们对鲎试剂的开发研究以及对鲎试验方法的不断认识,其技术越来越成熟,在1973年,美国食品药物管理局(FDA)承认鲎试剂是一种生物制品。1980年,美国药典第2版(uspxx)首先收载了细菌内毒素检查法(鲎试验法),随后,英、意、德、欧洲及日本药局方都相继收载了这一检查方法。同时其应用范围和领域也越来越广泛。 我国对于鲎试剂和鲎试验方法开发研制是在70年代中期开始的,到80年代初期,由于当时研究水平有限,加之缺乏标准化研究的基础,致使上千批次的实验结果无法统计分析。到了1983年卫生部授权中国药品生物制品检定所组织全国范围的大输液及放射性药品鲎试验检查法,并允许4种注射剂药品使用细菌内毒素检查法初检。1993年中国药典1990版第二增补本正式收载细菌内毒素检查法,但未涉及任何品种正文。1995年,中国药典1995版二部附录收载经修订后的细菌内毒素检查法;B种药品正文规定了细菌内毒素检查项。取消了原热原检查。
参数原理
2022.11.10

内毒素是怎么产生的?
自从内毒素被发现百年来,人们一直认为内毒素作为细菌细胞壁的结构成分只有在菌体死亡或人工裂解时才释放出来,但近年来发现革兰氏阴性菌在细菌对数生长期或细菌营养缺乏时也释放内毒素。 应用电子显微镜技术发现,在细菌对数生长期,革兰氏阴性菌外膜过度合成,过剩的外膜在菌体表面形成许多“疱疹样结构”( struture afblebs),内毒素以“出疱疹”的方式被释放到周围介质中去.菌体外膜上的“疱疹”有大小两种类型,“大疱疹”在菌体分裂产生横隔的过程中形成,随菌体分裂成两个细菌而死亡;“小疱疹”存在于菌体的外表面,在细菌生长过程中不断产生,随时释放到外环境中。尽管这种以游离状态存在的内毒素仅占整个细菌内毒素的一部分,但值得注意的是这种持续释放的内毒素可能在其对宿主的反应中发挥一定重要作用。
参数原理
2022.11.10

内毒素检测的方法有哪些?
内毒素的检测有传统的家兔热原试验(rabbit pyrogentest,RT)、鲎试验法(limulus test,LT)以及其他的一些特殊方法。 (一)家兔热原试验法此方法即将一定量的被检标本静脉注人家兔体内,观察其注射后的发热情况,以决定所检标本中有无热原存在。RT法为一种定性检测内毒素的方法,应用历史较久,此方法本身有许多限制,如家兔对内毒素在反应上有个体差异﹑敏感度不高、不能定量测出内毒素等。 (二)鲎试验法细菌内毒素检查法也称之谓鲎试验法(LT法)英文为Bacterial Endotoxins Test,它是在1964年由美国学者Levin和Bang所发现,并在1968年正式建立的一种定性或定量检测微量细菌内毒素的分析方法。由于其灵敏度高,特异性强而且操作简便快捷,目前在许多科研和应用单位均得到了突飞猛进的发展和推广应用。自此方法建立以来的30年间,其在细菌内毒素本质和方法学的研究方面更是不断进步和发展,加之各种新型试剂和新型仪器的开发,不仅在药学,而且在微生物学及临床检验等领域的开发应用更为广泛。 LT法又称鲎变形细胞溶解物试验(limulus amoebocytelysate,LAL),是目前检测内毒素最敏感的方法。它比RT法敏感10~100倍以上,可测出微量的内毒素(0.1 ~ 1ng/ml),用微量光电法及其他改进方法可测到微微克(pg/ml)的水平。而在RT法中,敏感性高的家兔需注射1.5ng/kg 才能引起发热,而敏感性低的却需要注射0.5 ~10μg/kg才能引起发热。LT法可广泛地用于革兰氏阴性菌感染的快速诊断,对患者的血液、尿液及脑脊液可进行直接的检查。此法亦可用于检查各种注射剂内是否有内毒素污染。
参数原理
2022.11.09

应有鲎试验法检测食品中的细菌内毒素含量
食品安全关系到每一个人的身体健康。如今,人们越来越重视食品的安全质量,食品若是受到细菌污染,就会随着细菌代谢的过程中产生细菌内毒素,即使在高温高压下,细菌内毒素也不易被破坏。若是人体大量食用则会产生急性中毒的症状,少量长时间食用就会引起慢性中毒,对人体极其有害。所以对正式投入市场的食品都需要进行细菌内毒素检测。 肉类、蛋、蛋制品、牛奶、饮料等的细菌内毒素主要都是由革兰氏阴性菌引起的。 1、肉制品的细菌内毒素检测在肉类中的革兰氏阴性菌,大部分都是假单胞菌。若是食物存放的环境潮湿且气温又高,变形杆菌就很容易在肉类的表面快速繁殖,导致肉类变质腐败。其他类型的革兰氏阴性菌包括副溶血弧等也能在肉类中快速繁殖增长,所以这些肉类制品一般都含有细菌内毒素。使用鲎试剂检测肉类的细菌内毒素含量,也能从侧面知道肉类被细菌内毒素污染的程度。 那如何应用鲎试验法检测肉类中的细菌内毒素呢?首先,要对对一定数量的肉类制品进行预处理,即把肉类的脂肪去掉;接着将肉类打碎,再离心沉淀;然后用清液对肉类进行细菌内毒素检测。用鲎试验法检测肉类的细菌内毒素,能检测出0.1ng的细菌内毒素含量,可以拿来当作食品卫生质量的一个标准。有研究人员提议,若用鲜牛肉经过1000倍稀释后,将鲎试验记为(-),就可以说明该肉类的质量的安全放心的。 2、蛋、蛋制品的细菌内毒素检测革兰氏阴性菌也很容易引起蛋类的变质。一般来说,变质后的蛋类以及蛋制品都会含有很高的细菌内毒素含量,变质的咸鸭蛋和皮蛋的细菌内毒素含量尤其多。使用鲎试剂检测蛋及蛋制品的细菌内毒素含量,就可以把细菌内素素含量高的这些蛋、蛋制品挑选出来。 3、奶制品及饮料的细菌内毒素检测在1976年的时候就有报道说用鲎试剂能检测出患有牛乳腺炎的牛的乳汁中的细菌内毒素含量。后来,陆续有人发发现牛奶样品中也能发现细菌内毒素,含量能达到30~130μg/ml,特别的放置在常温下24小时的牛奶,细菌内毒素的含量也会逐渐增多,甚至会增多到16倍。 一般市面上销售的饮料、啤酒等,都可用鲎试验法记作(-)。但是有些饮料是发酵制成的,也需要用鲎试验法进行细菌内毒素检测。
参数原理
2022.11.09

11月9日起丨科德角国际诚邀您参加《中国药典》药物制剂质量控制技术研讨线上培训
为进一步强化药品生产全过程管理理念,帮助行业掌握《中国药典》相关技术要求,加强对药品生产用原辅包以及生产过程的控制,进一步保障药品质量,国家药典委员会将于2022年11月9日-11日在线上开展《中国药典》药物制剂质量控制技术研讨培训。科德角国际生物医学科技(北京)有限公司作为受邀企业,将由科德角国际的资深技术总监范玉明老师出席此次研讨培训活动。科德角国际技术总监范玉明老师的此次培训内容以《光度法细菌内毒素检测技术应用》为主题,围绕热原检测与其替代方法发展史、光度法细菌内毒素检测趋势、细菌内毒素检测应用质量控制等多方面内容进行学术报告及研讨,分享细菌内毒素检测领域的新进展、新理念、新技术、新成果。培训日程如下: 科德角国际始终专注于细菌内毒素检测领域,以推动行业优质高效发展为己任,诚邀您莅临此次研讨培训会进行指导、探讨与交流,以便我们能够为您和行业做出更好的产品,提供更优的服务。 直播及回放:请登录国家药典委员会网站,【公众服务】-【在线培训平台】,或下面网址观看:https://apphun4a61i1930.pc.xiaoe-tech.com
企业动态
2022.11.09

内毒素对血液循环系统的影响及机理
内毒素可引起白细胞和血小板减少,激活凝血、纤溶系统,产生出血倾向,可导致弥漫性血管内凝血(DIC)的发生。其机理在于内毒素(脂多糖)可激活Hageman(Ⅻ)因子,内毒素的活性成分类脂A与Ⅻ因子结合后,在白细胞参与下促使凝血酶的生成、使纤维蛋白原转变为纤维蛋白,从而导致弥漫性血管内凝血的发生。DIC的发生是通过Ⅻ因子启动内、外凝血系统引发的全身性症状。DIC严重时,可导致严重的休克症状。 1、内毒素(脂多糖)对中性粒细胞的作用及机理:内毒素(脂多糖)能引起人体血液中中性粒细胞的增多,这是人体对内毒素反应最敏感的指标之一,亚致热量(Subpyrogenicdose)即可引起中性粒细胞的减少,并伴之以粘连性增加,数小时后,中性粒细胞明显增多。原因是内毒素(脂多糖)初进入机体后,大量的中性粒细胞隐伏在毛细血管管床内;然后能诱释中性粒细胞释放因子(neutrophil releelsing factor)。促使中性粒细胞从骨髓释放至血流当中,脂多糖也能增强中性粒细胞的吞噬和杀菌能力,激活补体系统使之产生中性粒细胞趋化因子。促使大量中性粒细胞杀伤和吞噬病原菌(G菌),增强人体非特异性免疫功能,导致机体病变。但伤寒菌内毒素例外,始终使血循环中的白细胞数减少,机制尚不清楚。 2、脂多糖对血小板的影响:对实验动物注射足量的脂多糖后,均能导致动物血液中血小板明显减少,这是由于动物血小板膜上有脂多糖的免疫粘连受体,故易与脂多糖发生结合反应,而人和灵长类动物体中缺乏这种结构,所以脂多糖注入人体后,一般不引起血小板减少。但在大剂量注入后,也可引起血小板减少。 3、脂多糖对淋巴细胞的作用:脂多糖作用于B淋巴细胞,可增强B淋巴细胞DNA的合成,有文献报道:脂多糖是B淋巴细胞的非特异性有丝分裂源,可促进B淋巴细胞的分裂增值。脂多糖的促分裂活性存在于类脂A,而免疫原活性与脂多糖部分有关。所以说,脂多糖能提高人体体液免疫功能,革兰氏阴性菌感染后引发的机体发热是人体自我防御功能的表现,是积极的。 4、脂多糖对单核细胞和巨噬细胞的作用:单核细胞和巨噬细胞对脂多糖很敏感,仅ng量就可引起反应,脂多糖作用于巨噬细胞和单核细胞后,能促使细胞内非溶酶体酶(如乳酸脱氢酶、亮氨酸萘胺酶)和酸性磷酸酶、β-N-乙酰葡萄胺酶等溶酶体酶的增加,增强吞噬和巨噬细胞的杀菌能力。同时,脂多糖作用于巨噬细胞和单核细胞后,能产生多种中介物质,参与外源性凝血过程,脂多糖能刺激单核细胞和巨噬细胞产生集落刺激因子(GSF)。 5、对红细胞和血清铁的影响:脂多糖能抑制红细胞的生成,脂多糖直接作用于骨髓造血干细胞,抑制骨髓的造血功能。也有人认为沙门氏菌内毒素进入机体后,与红细胞膜形成复合抗原,激活补体系统,发生变态反应凝血(Ⅳ型)反应。内毒素可导致血清铁浓度的降低,所以说,内毒素血症的另一病症就是缺铁性贫血。 6、内毒素可经C3旁路或经典途径激活补体系统:脂多糖激活补体的途径有三条,即①经典途径;②替代途径;③不依赖抗体的经典途径。激活补体后,最终导致血压下降和血液动力学的改变,出现局部或全身性 shwartzman(炎症症状),并伴有轻微的发热症状。
参数原理
2022.11.08

内毒素对糖代谢的影响及其机理
内毒素直接或间接损害肝脏,引起糖代谢紊乱及酶学、蛋白代谢的改变。动物实验证明:内毒素(脂多糖)注入体内后初期可引起血糖的暂时升高,数小时后,可导致血糖的持续降低,表现为明显的低血糖症。 (1)致病机理:脂多糖能刺激肾上腺素的分泌和释放,使磷酸环化酶活性增强,加速糖分解,使血糖浓度表现为暂时性升高。 (2)低血糖症的主要机理:脂多糖进入机体数小时后,始作用于肝细胞,引起肝细胞的损伤,使糖原异生的关键酶(如葡萄糖-6-6磷酸酶;丙酮酸羧基酶peck;糖原合成酶)活性降低,抑制糖原的异生和分解,激活丙酮酸激酶,加速葡萄糖的分解氧化,并产生大量的热量,在临床上表现为发烧症或持续低血糖症。内毒素致肝脏枯否细胞功能受到抑制,进而影响对内毒素的消除及对胰岛素的分解代谢。
参数原理
2022.11.08

内毒素的致病机理简介
内毒素的致病机理下图简图表示。 1、致热性极微量(1 ~5ng/kg)的内毒素注射入人体足以引起健康成人2小时内产生发热反应,且维持4小时左右,而对于革兰氏阴性菌感染,慢性消耗性疾病及晚期肿瘤病人,引起发热反应的最低内毒素剂量可低至0.2~0.5ng。 内毒素可使人体和动物致热,人体对内毒素致热原敏感。0.0001~0.005μg/kg 伤寒沙门氏菌内毒素即可引起人体发热,而家兔发热量比人高10倍左右。也就是说,在正常情况下,人体内内毒素的正常阈值保持在0.001~0. 05ng/ml之间,一旦人体免疫力下降或遭受某种致病菌侵袭时,细菌便在机体内繁殖崩解,释放出大量的内毒素,当体内内毒素的浓度高于这个阈值时,即可启动内外源致热原引起机体发热。 内毒素引起发热反应的作用机理并不是直接的,而是通过诱生内原性热原(endogeneons pyrogens,EPS)所起的间接作用。内毒素作用于肝枯氏细胞、中性细胞、中性粒细胞等,使之释放内源性热原质,再刺激下丘脑体温调节中枢所致。 目前已知EPS主要有三类,即肿瘤坏死因子(tumor necrasis factor,TNF)、白细胞介素- 1(interlenkin 1,1L-1)、β2-干扰素(interonB2,IFN-Bz)。当内毒素激活单核/巨噬细胞后这些细胞可合成和释放IL-1。IL-1是分子量12000~18000d的糖蛋白,为一族多肽类,分为IL-1α、IL-1β两个亚型,在70℃时,30分钟即失去其致热性。给动物静脉注射重组的IL-10. 1 μg/kg 就可引起发热。发热呈现双峰型,第一个热峰在注射后1.5~2小时出现,第二个热峰在注射后2~4小时出现,已经证实IL-1在作用于下丘脑体温调节中枢的过程中,刺激PGE2的形成。TNF系由单核/巨噬细胞受内毒素(ET)刺激后释放出的一种分子量为17000d的蛋白质。小剂量(1μg/kg)TNF引起的发热为单峰热,给家兔静脉注射后的1小时达到高峰。大剂量TNF(> 10μg/kg)可引起双峰热,第二个峰值在注射后3~4小时出现。TNF能刺激下丘脑合成PGE:及诱导IL-1的产生。 实验表明:应用内毒素(ET)后双峰的第一个热峰是TNF直接作用于体温调节中枢而产生的,第二个热蜂则是由于IL-1作用而产生的。IFN-β2分子量约为15000d左右,由淋巴母细胞或致敏的淋巴细胞所产生。由IFN-β2引起的发热呈单相型,过程较慢,峰值在注射后2~3小时达到。IFN-β2可刺激下丘脑释放PCE,直接作用于体温调节中枢而引起发热。 内毒素及诱生内原性热原均属于大分子物质,难以通过血脑屏障,故认为诱生内原性热原引起发热是通过室周器官(cirumventricular organs,CVO’S)而到达下丘脑的。此处的毛细血管是由孔状连接的内皮细胞构成,而在脑的其他部位,毛细血管是由紧密连接的内皮细胞构成。这样大分子物质就可由CVO’S处进人脑组织。CVO’S通道的终点位于第三脑室的嘴侧壁,其一面紧靠视前区,目前认为此处是发热介质由体循环到达下丘脑体温调节中枢的重要通道。诱生内原性热原作用于体温调节中枢的温敏神经元,使体温的“调定点”上升而导致发热。 另有研究报道:内毒素可直接作用丘脑下部,刺激体温调节中枢,导致机体发热。
参数原理
2022.11.04
内毒素的生物学活性是怎么样的?
细菌内毒素即脂多糖(LPS)是由脂质和多糖组成的大分子物质,其活性中心为类脂A。多种生物学系统的实验证实:类脂A可介导内毒素几乎所有的生物学活性,LPS及其类脂A对机体可发挥广泛的生物学作用,如致机体发热、休克、弥漫性血管内凝血、B淋巴细胞分裂、内脏器官实质性损伤等,严重者可危急生命,常见的疾病如发热、感染性疾病,肠道疾病、肝衰、胰腺炎等都与内毒素在体内的存在密切相关,且其引起的死亡率甚高,美国每年因内毒素致死的病人达10万人之多。 一、内毒素生物学活性1、非特异地与细胞膜结合,干扰细胞膜正常功能。 2、破坏线粒体,影响ATP酶和氧化磷酸化的偶联过程,使能量代谢发生障碍:线粒体是细胞的“动力工厂”,其功能依赖于膜结构的完整。线粒体膜类脂含量高,并有一种双磷脂酰酯甘油酯的特殊成分,与LPS亲和力很强,它以酰基与类脂A结合,引起线粒体膜结构紊乱,类脂A置换出三联体头部的ATP酶和基底部的细胞呼吸链复合物I,使氢及电子的传递发生障碍,细胞丧失正常呼吸功能,加上类脂A破坏丙酮酸脱氢酶系功能,致使丙酮酸进入三羧酸循环受阻,NADH、ADP、乳酸等代谢产物堆积,造成胞内代谢酸中毒,增强了溶酶体酶活性及H2O2的毒性作用,加重了因ADP不足引起的细胞水肿,加速了细胞自溶死亡。 3、损伤溶酶体膜,造成细胞损害:进入机体的LPS被细胞吞饮后,在胞内形成吞噬体,再和初级溶酶体酶融合形成次级溶酶体。LPS便引起溶酶体膜的结构紊乱,使其破裂释放溶酶体酶于胞浆内,溶酶体酶作用于细胞自身的线粒体等细胞器,加速细胞死亡。溶酶体酶还可作用于细胞本身的胞核,促进特异性的mRNA转录,使胞内溶酶体水平大大提高,更加速了细胞自溶过程。 4、激活腺苷酸环化酶:腺苷酸环化酶镶嵌在细胞膜的磷脂双层中,由受体、传递链、催化点三部分组成,受体在细胞膜表面和配体结合,信息经传递链激活膜内面的催化点,使ATP转化为cAMP。但类脂A却绕过受体直接和传递链结合,激活催化点,使胞内cAMP浓度增高,cAMP作为第二信使再激活cAMP依赖性蛋白激酶,在不同细胞中产生不同效应,如LPS作用于肠上皮细胞的腺苷酸环化酶,促进液体分泌引起腹泻。 5、对微粒体的影响:人的肝和肾上腺细胞中有大量微粒体,含有丰富的混合功能氧化酶,能直接利用氧分子参与氧化还原反应,分别和药物﹑激素的分解代谢以及类固醇激素的合成密切相关。混合功能氧化酶以辅酶Ⅱ为供氢体、细胞色素P-450为电子传递体。内毒素将细胞色素P-450中的血红素解离出来,使其失去电子传递功能,同时内毒素激活微粒体中的血红素加氧酶,迅速分解血红素基因。这可能是革兰氏阴性菌菌血症患者肾上腺皮质功能明显降低的直接原因,另外肿瘤病人在接受化疗过程中,免疫力遭到严重抑制,易并发革兰氏阴性菌感染,于是内毒素随血流到肝脏,破坏肝脏的解毒能力,使某些化疗药物半衰期明显延长,毒性作用增强,加速病人的衰竭死亡。 6、改变机体的免疫功能,包括刺激巨噬细胞,促使B淋巴细胞有丝分裂,诱导干扰素,并有佐剂作用和抗肿瘤作用。细菌内毒素及类脂A诱导的其他生物学作用见表1。
参数原理
2022.11.04

使用鲎试验法检测水质内毒素含量的优点
水质卫生安全的细菌学指标的主要标准一般以大肠菌数指数和大肠菌数值为主。虽然这些方法使用的原料简单,操作起来也方便,但每一个标本都需进行检查,而检查结果也需要24至48小时以内才会出结果,而使用鲎试验法检测水质卫生,在取样后2小时内便能得到结果,速度非常快。 据国外报道,水里面的总内毒素的浓度(特别是结合内毒素浓度)与细菌菌落数呈正比关系。如果以细菌菌落对总内毒素或结合内毒素的对数换算值绘制出回归曲线,可以显示两者之间的高度相关性。因此,可以通过检测水中内毒素的量来了解水中的细菌污染程度。鲎试验法可以检测到每毫升少至100个革兰氏阴性菌,因此它是一种非常敏感的方法。此外,鲎试验法还可以检测管道水中的其他细菌,如嗜铁细菌、硫酸盐等。虽然这些细菌是非致病性的,但它们会引起腐败和难闻的气味,因此它们也具有一定的公共卫生意义。 Diluzio曾在8个城市采集的18份饮用水样本中,就测出16份饮用水样本中含内毒素,部分自来水内毒素可达800μg/ml。他还发现,取自密西西比湖作为自来水的样品中内毒素含量约为400g/ml,而经过处理的自来水中的内毒素含量约为1μg/ml,这表明饮用水在净化过程中内毒素含量相应降低,但净化过程中的哪个阶段能起到这种作用不得而知。从两口井中采集的水样,一口为阴性,一口为阳性,可能与井深有关。推测深井水和土壤的过滤可以去除内毒素。 一般来说,鲎试验可以在没有稀释的水源上进行。对于污染严重的水源,离心沉淀后取上清液进行测定。
参数原理
2022.11.04

内毒素和淋巴细胞中的细胞膜非特异结合的介绍
内毒素是小鼠B淋巴细胞的致有丝分裂原,它能诱导B细胞增殖,并且分化成分泌抗体的浆细胞。T细胞的少数亚群也可以对内毒素起反应。这些作用的引发都是通过内毒素和淋巴细胞(特别是B细胞)上存在有内毒素受体,但是它也可以非特异地和内毒素结合。 已经知道,类脂A对淋巴细胞膜有一定的亲和性,碱水解的内毒素能增强这种亲和作用,而磷脂酰乙醇胺则能够抑制这种亲和作用。在大部分情况下,这种亲和作用表现为被动的内毒素非特异结合作用。内毒素和淋巴细胞膜也存在着主动的非特异结合,表现在内毒素可以显著降低细胞膜的膜电压,产生了3小时的后放电。这种现象在T细胞中没有被发现过。这种现象的产生依据是内毒素和淋巴细胞结合后钙离子通道被激活而引起的。 其他细胞:内毒素除了能够与以上所提到的红细胞、血小板、淋巴细胞发生非特异性结合以外,还能和许多哺乳动物的细胞种类非特异性结合,包括内皮细胞、巨噬细胞、肝细胞和白细胞,正常或经转化的成纤维母细胞等。内毒素和这些细胞的结合在大多数情况下,是被动的、有时也可以表现出主动结合的特性,如改变膜的流动性,破坏膜的完整性,改变其对Con A 的反应性等等,这些细胞的结合性质在很大程度上和红细胞、血小板以及淋巴细胞相类似,在作用方式和作用机理上也基本一致。
参数原理
2022.11.03
内毒素和红细胞及血小板细胞膜的非特异结合
1、红细胞 早在50年代,人们就发现如果将内毒素加入绵羊红细胞中能使红细胞被动致敏,产生凝集,并在外源性同种内毒素抗体存在下产生补体介导的细胞溶解反应,在红细胞膜肯定存在一种能识别内毒素并能与之结合的活性物质,现在我们知道红细胞对内毒素识别作用不仅可以通过特异性识别(内毒素受体),而且可以通过非特异的结合来完成。 多粘菌素B(polymyxin B)能和内毒素和活性成分类脂A特异结合。实验中发现,在多粘菌素B存在的情况下,红细胞许多依赖类脂A的免疫学和免疫病理学以及毒性作用被抑制了。这强烈提示类脂A能和红细胞表面的某种成分出现一种结合作用,而多粘菌素B则抑制了这种结合从而类脂A不能介导其相应的生物学作用。 内毒素和红细胞的非特异性结合不仅可以通过被动的疏水作用,而且可以通过主动结合的方式完成。 总之,内毒素和红细胞之间存在着非特异性的和特异性的结合,其中非特异性的结合可以通过主动的和被动的两种方式进行,碱处理过程的内毒素增加了这种非特异性的结合,改变了红细胞膜某些理化性质。 2、血小板 内毒素在血浆蛋白的存在下,能够影响血小板的某些功能。内毒素和血小板的结合是产生这种作用的基础。在大多数情况下,内毒素和血小板的结合还是通过类脂A和血小板膜磷脂的非特异结合完成的,这种结合具有相对比较低的亲和性,其原因可能是由于类脂A处于内毒素多糖成分之内,位于整个结构的中心,难以和血小板膜接近,不能充分结合。和红细胞相类似,类脂A能和血小板磷脂主动或是被动结合,其中大部分是属于被动结合,也存在着主动结合。表现在碱处理过的内毒素能增加血小板颗粒成分的分泌以及促进凝血的磷脂成分——血小板因子3的产生,这种作用呈剂量依赖性,其作用机理是因为碱处理过的内毒素增加了和血小板膜的结合,并且导致了血小板膜磷脂分子的重新排列和分布。
参数原理
2022.11.03

内毒素和细胞膜磷脂成分的非特异结合
革兰氏阴性菌的内毒素能够引起许多细胞的生理或病理反应,这些细胞包括肝细胞、脂肪细胞、淋巴细胞、巨噬细胞、多形核白细胞、血小板以及内皮细胞等。这些细胞产生的反应都是通过内毒素和细胞膜结合而介导的,内毒素和细胞膜的结合可以分为特异性(通过受体)和非特异性(通过细胞膜脂类成分)两种,后一种结合往往以不饱和性和无配体特异性为特征的。 内毒素和细胞膜的非特异性结合可以通过许多细胞膜的活性成分来实现的,这些活性成分包括糖蛋白、糖磷脂和磷脂。事实上,这几种活性成分都已证实能和内毒素非特异结合,但是,通过对内毒素活性成分类脂A的研究,人们发现细胞膜磷脂成分在其中起着十分重要的作用。 内毒素位于革兰氏阴性菌的细胞璧外膜的外叶,在形态上,细胞壁外膜是一种典型的双层结构,而内毒素的存在呈不对称性,并可与其本身、细胞膜磷脂以及外膜蛋白成分发生相互作用。多数学者认为内毒素类脂A和外膜磷脂的相互作用是通过亲脂性的疏水作用来完成的。 至于内毒素和宿主细胞磷脂的结合,从结构上看,可以分成主动和被动的结合两种。主动结合是指磷脂和内毒素结合以后,磷脂部分破坏了内毒素大分子的三级结构,或者是内毒素部分破坏了磷脂的三级结构,最后形成了一种新的内毒素-磷脂复合物,呈颗粒状构型。被动结合是指内毒素通过表面张力和/或疏水作用而吸附到细胞膜的磷脂颗粒上。 无论是主动或者是被动的结合作用,内毒素在磷脂结合的基础上,都能改变特定磷脂颗粒的物理和化学性质(如膜电压、膜完整性和流动性等),通过信号的转膜传递,影响细胞的状态和功能,至于这两种方式的选择,则主要取决于内毒素的类型(光滑型或粗糙型)疏水表面的理化性质等等。
参数原理
2022.11.03

菌苗、疫苗、血液制品及干扰素,如何检测细菌内毒素?
生物制剂的制备过程中也需要检测细菌内毒素。生物制剂的种类很多,其中包括菌苗、疫苗、血液制品及干扰素(IFN)等那它们如何检测细菌内毒素呢? 一、菌苗菌苗在制备时,多多少少都会受到内毒素的污染,尤其是革兰氏阴性菌菌苗,受内毒素污染的程度会更高。为了能更好地检测菌苗中的细菌内毒素,可以采用灵敏度高、检测速度快且准确并可定量的鲎试验法。有试验检测发现,用鲎试验法检测多糖体菌苗的细菌内毒素含量时,内毒素含量程度最高的菌苗(14000ng内毒素/100μg)其发热反应率也是最高的,1.8~1.9%的肛温都超过38.5℃;但细菌内毒素含量低的菌苗(600ng/100μg),反应情况也较低。根据这个试验可以知道,检测细菌内毒素使用鲎试验法,能有效对菌苗生产技术进行改进,其反应率也会降低,从而能让菌苗达到满意的提纯效果,所以鲎试验法是菌苗检测细菌内毒素的一个不错的方法。 二、疫苗现今,鲎试验法的普及程度已经很高,且在试验中都是高效可靠的。鲎试验法也被成功用在流感疫苗的质量控制上,用于检测流感疫苗中的细菌内毒素含量。有研究人员从美国6家厂家生产的26批流感疫苗进行细菌内毒素检测,结果发现这次流感疫苗的批间的细菌内毒素含量的差别非常大,所以美国FDA特制了一批标准流感疫苗用于安全试验检测使用,要求凡是新生产的且用于人群免疫使用的A/swine株流感疫苗的细菌内毒素含量不能超6ng/ ml。这个规定大大提高了A/swine株流感疫苗的质量。 三、血液制剂鲎试验法也被用于血液制品的质量控制当中。因为血浆或血清中存在了内毒素抑制蛋白,所以用鲎试验法对血液制剂进行细菌内毒素检测时,必须要先对血液制剂进行预处理。使用鲎试验法检测血液制剂的细菌内毒素含量,可以高效地杜绝被细菌内毒素污染的可能性,能有效保证血液制剂的安全性。 三、干扰素(IFN)淋巴细胞在体外经诱导剂刺激后会产出一种醣蛋白,这就是我们常见的干扰素(IFN)。越来越多的研究发现,干扰素(IFN)会引起发热、刺激B淋巴细胞、抑制T淋巴细胞等症状,和细菌内毒素有很多一样的反应。但是部分人认为之所以会出现和细菌内毒素一样的症状是因为干扰素(IFN)在制备过程中受到了细菌内毒素的污染。所以使用鲎试验法检测干扰素(IFN)的细菌内毒素含量,能有效降低干扰素(IFN)的副反应症状。 总而言之,使用鲎试剂法检测生物制剂的细菌内毒素含量,能更好的保证生物制剂的使用安全。
参数原理
2022.11.02
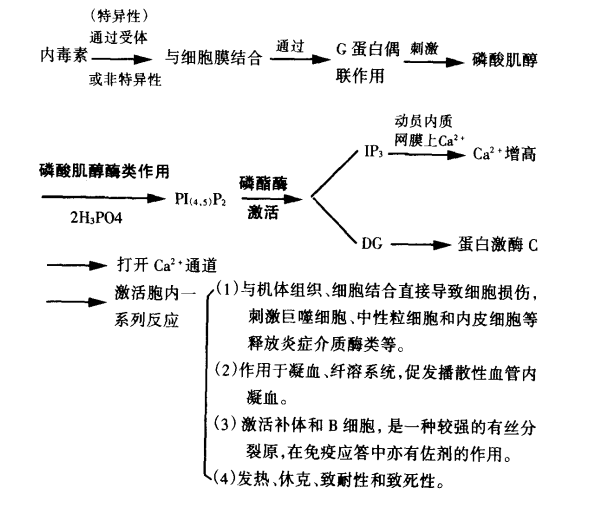
脂多糖结合蛋白介导细胞激活的信号传递是怎么样的?
脂多糖(LPS)作用的靶细胞主要是单核/巨噬细胞。LPS在诱导单核/巨噬细胞合成的分泌细胞因子的过程中脂多糖结合蛋白(LBP)起着信息转导的作用。 血浆中的 LBP与LPS的类脂A部分连接形成高亲和性复合物,这一复合物再经膜蛋白CD14受体连接于巨噬细胞表面,使巨噬细胞快速激活,并诱发肿瘤坏死因子(TNF)的产生及白细胞介素-1β(IL-1β)mRNA 的转录。单核/巨噬细胞在接受TNF、LPS、IL-1刺激后释放IL-6,LPS与TNF、IL-1、干扰素r(IFNr)之间具有协同作用。IL-6一方面转导TNF、IL-1的作用,另一方面又下调TNF、IL- 1的表达,起负反馈调节作用,因此,TNF、IL-1、IL-6可能单独或联合作用参与内毒素中毒的病理过程。 何炬斌(1995)认为内毒素和细胞膜结合后将信号传递入细胞内释放内源性介质完成信号的转膜传递。内毒素和细胞膜结合之后,通过G蛋白的偶联刺激细胞内磷酸肌醇系统的周转,产生第二信使——三磷酸肌酸和 甘油二酯,并通过它们引起胞内Ca2+浓度变化及激活PKC(蛋白激酶)。在 PKC的催化下,细胞内的功能蛋白完成磷酸化、最后实现一系列的细胞反应。
参数原理
2022.11.02

细胞识别内毒素机制以及CD14和其他一些脂多糖的识别分子的介绍
一、细胞识别内毒素机制脂多糖(LPS)与脂多糖结合蛋白(LBP)形成复合物后,能有效地与可溶性CD14(sCD14)或膜结合CD14(mCD14)受体结合,并活化细胞反应。LPS在细胞膜上的识别在LPS的摄取和诱发穿膜信号乃至细胞的活化是必须的。虽然有几种蛋白质与此有关,但其中CD14提呈LPS至具有功能性的LPS受体上。sCD14途径控制内皮细胞和上皮细胞对LPS的反应。最近研究表明,通过依赖LBP/CD14途径的LPS诱导细胞活化过程中,细胞中蛋白酪氨酸磷酸化起重要作用。 二、CD14和其他一些脂多糖的识别分子脂多糖(LPS)或内毒素可激发体内广泛的,非特异性连锁反应导致多种潜在介质的分泌和通过激活巨噬细胞和单细胞而产生一些细胞因子,这些细胞因子(如IL-1、IL-6或IL-8或TNF-α)的大量产生,促成了内毒素休克的病理生理过程。近年来,人们在内毒素识别和细胞反应启动的机理研究中发现了一组重要的蛋白质分子,它们分别为脂多糖结合蛋白(LBP)和脂多糖受体簇分化抗原14(CD14)。体外实验证实,LBP/CD14系统能明显提高多种细胞对内毒素的敏感性,可使内毒素活性提高数百倍至数千倍。 1、LPS结合蛋白(LPS binding protein LBP)LBP是一种糖蛋白,分子量为58~60kd,主要由肝脏合成释放入血。1986年Tobiss首次从兔急性反应血清中分离纯化出脂多糖结合蛋白,正常人和许多动物血清中都有LBP存在,但一般浓度低于0.5ng/ml。只有在应激状态下,其浓度可高达50ug/ml。LBP能识别LPS类脂A部分,与LPS有很高的亲和性,并且能与带有LPS 的菌体或粒子结合,具有促进这些粒子与巨噬细胞(MΦ)结合的调理作用。在细胞对脂多糖(LPS)的应答过程中,LBP主要起催化作用,即LBP将LPS单体从其多聚体中转运到CD14,加速LPS与 CD14的结合,1个分子LBP可促使上百分子LPS与CD14结合。实验研究表明,当LPS浓度为0.01 ~ 1ng/ml 时不能单独诱导MΦ表达TNF,但如果有LBP存在,则可诱导MΦ表达TNF。单核/巨噬细胞表面存在针对IPS-LBP复合体的受体。LBP的生物学功能主要包括:(1)与LPS形成复合物促使机体更有效地探测到小量的LPS;(2)促进LPS-CD14复合物的形成;(3)结合革兰氏阴性杆菌,增强吞噬作用,起到调理素功能;(4)应用抗LBP抗体,中和血清中的LBP以减轻LPS攻击机体引起的一系列反应。 2、CD14CD14是细胞膜蛋白家庭成员之一,一般认为,CD14以两种形式存在,即膜结合CD14(mCD14)和可溶性CD14(sCD14)。mCD14是分子量为55kd的糖蛋自,以糖基磷脂酰肌醇(glycosylphosphatidy linositol GPI)锚形物附着在单核巨噬细胞和中性粒细胞表面。LPS-LBP复合物与细胞表面的mCD14结合,从而激活单核细胞或中性粒细胞。用磷脂酶处理单核-巨噬细胞,可以洗脱细胞表面95%以上mCD14。CD14不仅存在于膜表面,还可作为可溶性分子自膜表面游离至血清或体外培养的单核细胞上清液中(sCD14)。sCD14是含有链内二硫键的单链蛋白质,完整的重组sCD14(rsCD14)含有348个氨基酸,rsCD14,氨基末端152个氨基酸是其活性部分,可能结合LPS并介导其生物活性。其中,氨基末端57~64位氨基酸是LPS的结合部位,7~10位氨基酸与信号传导有关。sCD14存在于血清,尿液和 mCD14阳性细胞的培养上清液中,无GPI结构,正常血清中sCD14约为4ug/ml。sCD14分子有两种形式:49kd和55kd,但正常人血清中只有49kd的sCD14。有关sCD14的生物学作用至今尚不明了,sCD14最初被认为是体液系统中LPS的抑制物质,推测巨噬细胞释放的sCD14可以起到保护作用,并设想重组sCD14(rsCD14)应用于革兰氏阴性菌败血症的治疗中,但研究表明sCD14的生物学作用与其浓度密切相关。sCD14须达其正常血清浓度的10倍才能起到抑制LPS毒性的作用。
参数原理
2022.11.02
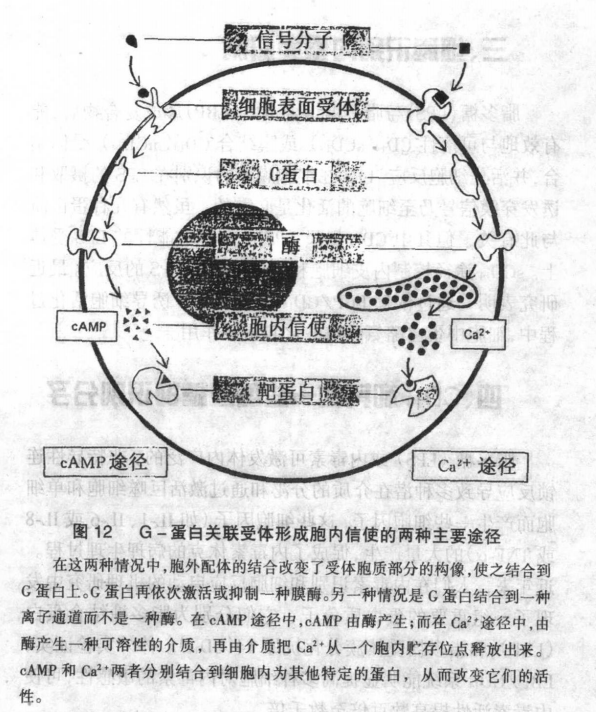
内毒素细胞内受体及其介导的信号传递
受体是细胞表面或胞内亚细胞组分中的一种蛋白分子,它可以识别并特异地与有生物活性的化学信号分子(配体)结合,从而激活或启动一系列生物化学反应,使该化学信号分子发挥其特定的生物效应。水溶性信号分子通过细胞表面受体起作用,而脂溶性信号分子则主要通过细胞内受体行使其效应。 目前,已知至少有三类细胞表面的受体:通道关联受体(channel-linked receptor)、催化受体(catalytic receptor)和G-蛋白关联受体(G-protein-linked receptor)。这是按其转导信号的不同机制而分的。 (1)通道关联受体:通道关联受体是一种递质闸门离子通道(transmitter-gatedion channel),它主要执行电兴奋细胞间快速的突触信号传递。 (2)催化受体:当催化受体被激活时,可作为酶直接起催化作用,几乎所有已知的催化受体都是跨膜蛋白,在细胞质的部分是一个有酪氨酸特异性的蛋白质激酶。 (3)G-蛋白关联受体:G-蛋白关联受体可以间接激活或失活一个不相连的膜结合酶或离子通道。受体与酶或离子通道之间的相互作用是通过第三种蛋白,称为CTP结合调节蛋白(G蛋白)来完成的。G-蛋白关联受体通常激活一系列反应,改变一种或几种小的细胞内信号分子的浓度,这些细胞内信号分子亦称为胞内信使(intracellular messengers)或胞内介质( intracellular mediators),这些胞内信使,随后起作用,依次改变胞内其他靶蛋白的行为。 主要的胞内信使(intracellular messeger)(亦称第二信使)包括环腺苷酸(cAMP),环鸟苷酸(cGMP),钙离子(Ca2+),1,4,5-三磷酸肌醇( InsP3)及 1,2-二酰基甘油(DG)(图11)。 其中cAMP和Ca2+是两种最重要的胞内信使。 cAMP和Ca2+信号分子的产生途径不同,但两者都要G蛋白的参与,几乎所有动物细胞都使用这两种胞内信使作为信号分子(图12)。 (4)细胞激活过程中的信号传递:细胞外的信号是通过何种途径影响细胞内物质的代谢和细胞的功能,这是当前细胞生物学研究的核心问题。现已知,细胞激活过程中的信号传递过程为:细胞外的信号(配体)可能和细胞膜上的特异性受体相结合,通过G蛋白系统偶联,激活磷脂酶C,产生细胞内第二信使——三磷酸肌醇(IP3)和二脂甘油(DG),从而使细胞内Ca2+离子浓度上升以及蛋白激酶C被激活,最后生成花生四烯酸类产物以及细胞内蛋白质的磷酸化,完成整个信号传递的过程。 内毒素激活细胞的过程大致上同上述信号过程一样,内毒素可以和许多种类细胞的细胞膜产生特异的或非特异的结合,在某些细胞膜上已成功分离出了内毒素受体,如脂多糖结合蛋白/脂多糖受体(LBP/CD14)。但多数情况内毒素和细胞膜的结合是非特异的,这种结合可能是内毒素的脂质成分——类脂A和细胞膜上磷脂成分通过疏水作用被动结合,也可以主动结合,这两种结合都可以改变细胞膜的某些理化性质。
参数原理
2022.11.01
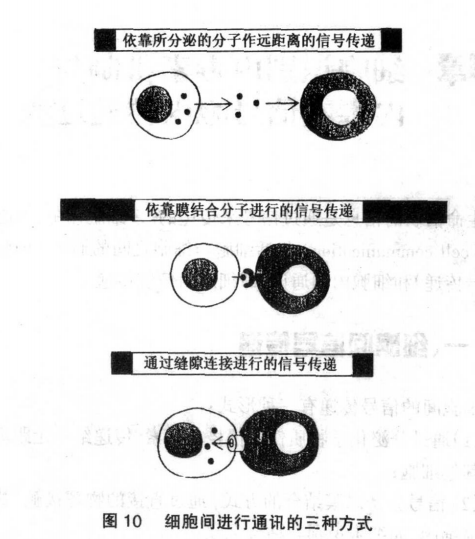
内毒素细胞间的信号传递是怎么样的?
生命活动的信息是通过信号来传递的。多细胞体中,细胞通讯(cell communication)包括细胞与细胞之间的通讯(即胞间信号传递)和细胞内的通讯(即细胞内信使传递)。 细胞间信号传递 细胞间的信号传递有三种形式:(1)通过分泌化学物质作为信号如激素,传递给一定距离外的其他细胞;(2)信号分子以膜结合的方式,通过直接的物理接触,影响其他细胞,如细胞识别与粘合现象;(3)细胞间形成间隙连接,小分子物质可通过间隙连接直接进入另一个细胞的胞浆内并发生作用,从通讯的联络方式看,第一种形式属于间接联络类型,第二三种形式属于直接联络类型(图10)。
参数原理
2022.11.01
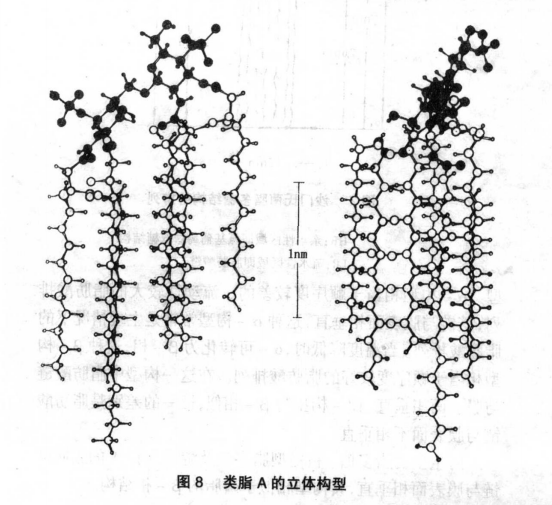
细菌内毒素中类脂A及脂多糖的立体构型介绍
在《细菌内毒素中脂多糖之类脂A化学结构的简介》一文中有提到:类脂A(又称脂质A),它是脂多糖的毒性及生物学活性中心。为一种糖磷脂,现认为细菌外膜的外层可能是类脂A。接下来介绍:类脂A及脂多糖的立体构型。 一、离体脂多糖的双层结构应用X-衍射技术已经证明脂多糖无论处于干燥状态还是在水溶液中,均能形成双层结构。根据脂多糖的来源不同,它们可形成不同厚度的双层结构。例如奇异变形杆菌脂多糖可形成9.6nm厚度的双层结构,大肠杆菌B/r脂多糖可形成10nm厚度的双层结构(根据脂多糖中的含水量不同其厚度可有改变)。高度纯化的明尼苏达光滑型脂多糖,粗糙型脂多糖及类脂A 分别形成11.2,6.0及5.2nm的双层结构。根据这些数据可以算出脂多糖或类脂A单层结构的厚度(上述数据各除以2)。因而:(1)类脂A的厚度约为2.6nm(=5.2/2nm)。(2)粗糙型脂多糖的厚度相当于3.0nm,亦就是说由KDO形成的厚度约为0.4nm。(3)光滑型脂多糖的厚度约为5.6nm,亦即由脂多糖中多糖部分(包括KDO(核心寡聚糖))所形成的厚度约为3nm。由于脂多糖的O-多糖链部分可携带20 ~40或更多个重复寡聚糖单位,故而如果以伸展形式存在的话,其长度可超过10nm。因而上述结构提示脂多糖的多糖部分以卷曲的形式存在(图7)。 二、脂肪酸链的排列由于类脂A与磷酯具有相似的构造,故而它们在细菌外膜中的构型是可比的。根据温度、水含量与化学本质的不同,磷脂的脂肪酸可以不同的形式存在,最常见的α,β及β′构型。α-构型相当于顺序度较差的、流动性较大的脂肪酸排列,其链与膜表面相垂直,这种α-构型常常是生理情况下的脂肪酸排列。当温度降低时,α-可转化为β-相,这种β-构型相当于顺序度较好的脂肪酸排列。在这一构型中脂肪酸链与膜表面相垂直β′-构型与β-相似,唯一的差别是脂肪酸链与膜表面不相垂直。野生型的脂多糖、粗糙型脂多糖及游离类脂A的脂肪酸链与膜表面相垂直,其构型相似于磷脂的β-相结构。 三、类脂A的分子模型随着先进的生物物理及化学方法的建立,目前已有可能建立游离类脂A的立体分子模型。下列所讨论的类脂A分子模型,主要是依据X-衍射分析结果及α能量计算方法所取得的综合分析结果。类脂A的亲水性D-GlcN(2-脱氧-2氨基-葡萄吡喃糖)双糖骨架以一个平面存在,且与外膜形成一个45角。类脂A所含的脂肪酸均存在于这一平面的一侧并与膜表面成90°角存在,这一构型如图8所示。 四、存在于细菌外膜中脂多糖的构成脂多糖在外膜中的结构示意图如图9所示。这一结构显示出以下几个特点:(1)与天然磷脂比较,脂多糖显示出高度的有序性,这是由于其中存在的类脂A的高顺序度所决定的,这种形式的结构显得相对的僵直。(2)脂多糖的O-多糖链部分以高度螺旋的状态存在。(3)类脂A的亲水性骨架(双磷酸化的D-GlcN双糖)与外膜表面成45角。(4)类脂A的脂肪酸全部存在于膜的内叶,并形成相对独立的小区域。
参数原理
2022.11.01

细菌内毒素化学结构之O-特异多糖链和核心寡聚糖的介绍
内毒素的主要化学成分为脂多糖(LPS),脂多糖由结构及生物学活性互不相同的三部分组成,即O-特异多糖链(O-specific pelysaccharide chain)、核心多糖(Coreoligosaccharide)、类脂A(lpidA)。有的学者将脂多糖的化学结构分为两大部分,即多糖(polysaccharide)和类脂A(Lipid A),多糖部分又可进一步分为O-特异性多糖链及核心寡聚糖(图6)。 今天将介绍内毒素中脂多糖的O-特异多糖链(O-specific pelysaccharide chain)和核心寡聚糖。 1.O-特异多糖链O-特异性多糖链是脂多糖分子中最易变异的成分,位于脂多糖的最外层。各种不同属产生的脂多糖具有不同的О-多糖成分,同一种属细菌产生的О-特异多糖链其组成成分亦可表现出异质性,不同菌种特异性多糖链上单糖的种类排列顺序和多糖链的空间构型不同,构成O-特异性多糖链上抗原决定簇的不同,该抗原决定簇是各菌种血清学特异性的结构基础,在细菌分类鉴定,菌苗制备,免疫机制研究等方面有重要意义。一般讲O-特异性多糖链是由数个至数十个(最高达40个)低聚糖、3~5个单糖(重复单位)集聚而成的多糖链。常见的单糖有戊糖、4-氨基戊糖、已糖、α-氨基已糖、6-脱氧已糖等,例如大肠杆菌糖的化学组成为3、6-二脱氧-L-半乳糖;沙门氏菌具有甘露糖–鼠李糖-半乳糖组成的三糖基本骨架。 脂多糖所含重复单位的数目越多,则脂多糖О-多糖链亦就越长,分子量亦越大。重复单位的数目及链的长度取决于细菌的种类。即使是同一种细菌提取得到的同一批脂多糖,亦表现出很显著的异质性(О-多糖链可从0到具有50个以上的重复单位不等)。某些细菌О-多糖链可由寡聚糖单位重复联结而组成较长的链(枸橼酸杆菌,耶尔森氏菌等),有些细菌的О-多糖链则仅含有几个糖单位,或在某些情况下,O-多糖链完全缺乏,后者的脂多糖即为粗糙型-脂多糖(R-LPS)。 特异性多糖链决定脂多糖的O-特异性抗原性。革兰氏阴性细菌的О-抗原分类(血清学分类,亦即Kauffmann-White分类)即是根据这一结构进行的。目前已鉴定了100种以上脂多糖的O-特异性多糖链结构,包括许多从临床感染病人分离得到的阴性致病菌,并不断发现新的糖单位及某些特殊的糖类。 2.核心寡聚糖与O-特异性多糖链相反,脂多糖核心成分的变异甚小,位于类脂A 的外层。进一步可将核心分为外核(outer core)和内核(inner core)两部分。前者的组成一般含已糖成分,如Glc,Gal,GlcN等。而后者含有Heptose一些自然界并不常见的糖成分,如L-甘油型-庚糖(L-glycero-Heo-tose,Hep)和2-酮-3脱氧-D-甘醇型–辛酮糖酸(2-keto-deoxy-D-manno-octonate,KDO),庚糖及KDO是革兰氏阴性菌LPS所特有的多糖。同--属细菌的核心多糖相同,不同菌属之间的核心多糖各有差异,故有属特异性。
参数原理
2022.10.31

细菌内毒素中脂多糖之类脂A化学结构的简介
类脂A(又称脂质A),系脂多糖的毒性及生物学活性中心。为一种糖磷脂,现认为细菌外膜的外层可能是类脂A。各属细菌的类脂A 结构相似,无种属特异性,故不同革兰氏阴性细菌感染时,由内毒素引起的毒性作用十分相似。给实验动物注人类脂A时,可出现典型的内毒素反应,如发热、血液动力学改变、弥漫性血管内凝血及内毒素休克等。类脂A在革兰氏阴性菌感染的机理中亦起着十分重要的作用,故而类脂A可认为是细菌的一种致病因子。另外,类脂A参与细菌外膜的组成,在维持细胞壁的稳定性、屏障功能以及在细菌的生理学上亦起着十分重要的作用。 类脂A的化学结构类脂A通过α-糖苷键而直接连结于脂多糖内核的KDO分子上,这一糖苷键可经温和酸处理而发生裂解,由这种方法处理得到的物质称为游离类脂A(free lipid A),而存在于完整脂多糖分子中的脂质成分称结合类脂A(bound lipid A)。 随着先进的化学及物理学方法的建立,目前已弄清了许多革兰氏阴性菌类脂A的化学结构,特别应当提出的是,80年代以来,由于分析和化学合成专家的密切合作,已成功地合成了肠道菌(沙门氏菌和大肠杆菌及其他细菌)脂多糖的类脂A及类似物。这些研究对于了解类脂A结构与功能间的关系具有重要的意义。 一般来讲,典型的类脂A是由两个氨基葡萄糖(GlcN),以β-1,6-糖苷键相联而组成的基本骨架,双糖骨架的游离羟基及氨基可携带长链脂肪酸和磷酸集团,不同种属细菌合成的类脂A 的分子组成,以及其骨架上的取代基团均有差异,类脂A按结构可分为两类。 第一类:以沙门氏菌为例举例说明,沙门氏菌类脂A由D-葡萄糖( 一22%)、磷酸(一10% )及长链脂肪酸( - 60%)组成。分子量大约为2000d,以 B-1,6-糖苷键连接的氨基葡萄糖双糖构成类脂A骨架,双糖的氨基以酰胺键与β-羟基脂肪酸相连。而双糖上3,4,6-位上的羟基则以酯键分别与各种不饱和脂肪酸及D-3-羟基脂肪酸相连,脂肪酸链上的3-羟基有时可被碳元素数目相等的脂肪酸所酯化。 第二类:类脂A不含氨基葡萄糖,而是以2,3-二氨基-2,3-二脱氧-D-葡萄糖为骨架。糖分子中两个氨基被D-3-羟基四价癸酸酰化,含少量的脂肪酸。 第一类与第二类之间不发生免疫交叉反应。类脂A不溶于水,而溶于吡啶、三乙胺、二甲基亚矾及氢氧化钠等,类脂A具有免疫原性,注入动物体内可产生相应的抗体。 类脂A既含有疏水中心(酰化双糖),又含有亲水端(磷酸);既含有酸性基因,又含有碱性基因,因此它既是一个双相分子,又是两性分子,这就赋予类脂A特殊的功能和致病作用。它既能与多糖链相结合形成完整的内毒素分子——脂多糖,构成细菌细胞壁的重要成分,防止外界有害物质入侵,又能与宿主细胞膜的受体结舍发挥各种生物学效应。 类脂A可视作脂多糖的生物学活性“内毒素中心”,如其糖衍化物KDO能刺激人和小鼠单核细胞产生白细胞介素-1(IL-1),并影响小鼠巨噬细胞产生花生四烯酸盐代谢产物。当脂多糖由革兰氏阴性细菌表面释放后,可与一种或多种血清或血浆蛋白相互作用,然后结合至宿主细胞上的特异脂多糖受体上,引起前炎症介质产生。 至于究竟是释放出的脂多糖抑或是结合于细菌上的脂多糖表现对单核——巨噬细胞较强的刺激作用的问题,目前尚不清楚。有人仍对抗生素介导的内毒素释放能加重败血综合征的问题持有怀疑态度。因为不同抗生素诱发革兰氏阴性细菌脂多糖释放的能力各不相同,两种对细菌细胞璧起作用的抗生素的内毒素释放潜力也与呈菌血症小鼠的生存有所不同。
参数原理
2022.10.31
鲎试验法:最宜对药物进行细菌内毒素检测的方法
药物在被正式使用前,必须进行细菌内毒素检测,而检测细菌内毒素的方法,又属鲎试验法最为适宜。美国FDA已把鲎试验法定为检测药物细菌内毒素检测的法定方法。应当知道,在应用鲎试剂对药物进行细菌内毒素检测时,该鲎试剂的灵敏度的值不应低于0.2EU/mL,且同时应当知道药物中细菌内毒素的耐受度限量值。 若是检测的药物没能通过鲎试验法却通过了家兔法,这种药物是不能被批准使用的,但该家兔法由官方进行测试的除外。若已知该药物对鲎试验法有抑制或者增强反应,则应当对该药物进行抑制或者增强试验。 检测抗生素的细菌内毒素含量最宜使用鲎试验法。这是因为大多部分抗生素对鲎试剂并没有抑制反应,可以对这些抗生素应用鲎试验法检测细菌内毒素。但也有部分抗生素溶液对鲎试剂有抑制反应,所以进行抑制试验时必要的。可以用无热原水进行稀释,知道该抗生素溶液对鲎试剂没有抑制反应才能进行试验。 出现最典型抑制鲎试剂反应的药物要属青霉素这一类药物,但是对于该药物的抑制原理并不明确。有些人认为青霉素出现抑制作用是因为该药物的细菌细胞壁有酶;也有人说青霉素的抑制作用和血清蛋白结合率有关,但是最终都没有一个最确切的定论。但是必须知道的是:检测抗生素的细菌内毒素含量,鲎试验法比家兔法灵敏10~100倍。 使用肿瘤化学疗剂中的副作用与应用内毒素的反应是很像的,所以肿瘤化学疗剂可能是因为内毒素污染了才会出现副作用。所以,对药物进行细菌内毒素检测是很有必要的,让药物中的细菌内毒素含量控制在安全水平以下,来减少不良反应。部分肿瘤化学疗剂存在细胞毒性,所以家兔法不适合使用,但是鲎试验法仍然是最适宜的
参数原理
2022.10.31
