(1)细胞核的分离: 将培养的细胞制成细胞混悬液,或以胰蛋白酶消化法于培养盖上收集培养细胞。应用MgSO4染色分离方法分离细胞核(Van den Engh et al 1986, 在Traok和Van Den Engh 34章 )。细胞混悬液浓度为5×106/ml。利用RNA酶消化后,使核从细胞分离,细胞核混悬液浓度为4~5×106细胞核/ml。(2)细胞固定和酸的处理:在5 ml试管内加冷的100%酒精不断旋转以达到满意的固定。在冰上停留10 min。在4 ℃离心(×150 g)10 min。重复加三次冷的100%酒精入试管内,离心,倾去。置于冰上10 min,再离心。然后加入相当核悬液1/2量的0.1 n HCl , 0.5% Triton X-100。室温停留10 min。加入IBM-0.25%Triton X-100(IBM配方:50 mmol/l KCl, 10 mmol/L MgSO4, 5 mmol/L HEPES pH8.0)。再离心,重复IBM漂洗(这时细胞核可在不染色情况下,以荧光显微镜观察)后,以2×SSC-0.1%Tween漂洗1×3min,继之加入等量2%的多聚甲醛在1×PBS-5 mmol/l MgSO4。在室温静置站立10min。倾去上清液,加IBm -Triton X-100漂洗,离心,使细胞核混悬液最终浓度为108/ml,(可用IBM-Triton X-100稀释约50倍,在血球计数器计数),混悬液镜检应含单个,完整的细胞核。(3)细胞核混悬液杂交①配制杂交混合液:甲酰胺5份,20×SSC1份,50%硫酸葡聚糖2份,pH调至7.0。此原液(stock solution)可贮存在4 ℃冰箱内。应用时加1份10 mg/ml鲱鱼精子DNA(herring sperm DNA)。②混合1 μl的细胞核混悬液(108/ml)与18μl的杂交混合液,充分混匀。将此19 μl混合液移入1.5 ml容积的Eppendorf 管中(核含量约为105)。③加入100 ng/每管的AAF标记DNA探针(如为生物素标记DNA探针浓度为20~40 ng/每管)。④置70 ℃10min使DNA探针和核DNA变性。⑤和组织切片与DNA探针杂交方法相较,不同的是在加热变性后切勿置冰上迅速冷却以终止反应,而应迅速转入37 ℃孵育过夜。(4)杂交后漂洗在每管中加入1.25 ml 50%甲酰胺-2×SSC(pH7.0),在42 ℃静置10~15min。偶尔旋转以助混匀。冷却至室温。加100 μl经dimethylsuberimidate(DEMS)处理的血细胞(107/ml)混匀,离心,室温,10min,轻弹试管使沉淀的小块散开,加入1.25 ml 2×SSC(pH7.0),42 ℃,继之,静置于室温10~15min,如前离心,再加1.25 ml IBM-Triton X-100,室温静置5min,离心。注:DEMS处理红细胞方法:经漂洗并离心去除白细胞和血清的红细胞在盐液如PBS中,细胞含量为108/ml,以K2CO3和DEMS溶液处理3次,第1次:K2CO3为20 mmol/L,DEMS为3 mmol/L,以后2次:K2CO3依然为20 mmol/L,而DEMS为10 mmol/L。在应用前将K2CO3和DEMS液混合加入红细胞混悬液中。在最后2次漂洗液中,应用100 mmol/l K2CO3将pH调至9~10。在25 ℃,15min后,加入50 μl,100 mmol/l 的柠檬酸(citric acid)/每ml细胞混悬液的浓度以达固定红细胞的目的。固定的红细胞离心倾去上清液后,用2×SSC稀释到108/ml,加0.1%叠氮钠可在4 ℃保存至少1年。(5)AFF标记的荧光显示:加200 μl的PBS含0.05%Tween和2%正常血清(NGS),轻轻振荡混匀,室温静置10min,加20 μl 1:100的单克隆抗AFF抗体,37 ℃孵育45min,加1.25 ml的PBS-Tween,室温静置10min,加20间歇性振荡,离心,倾去上清液,加200 μlPBS含0.05%Tween –2%NGS,振荡,室温静置10min,加20 μl的羊抗小鼠–FITC荧光标记抗血清,稀释度1:100~1:300。孵育于37 ℃ 45min,加1.25 ml PBS –Tween,室温静置10min,离心,倾去上清液。(6)生物素标记探针的荧光显示:加200 μl 4×SSC含0.1%Trion X-100 和5%BSA。室温静置10 min后,加20 μl抗生物素标记FITC抗血清15 μg/ml,孵育在37 ℃ 30min,以1.5 ml 4×SSc –0.1% Trion X-100洗1次,加入1.25 ml IBm –Triton X-100,室温静置10~15min,间歇振荡、离心。(7)荧光显微镜观察:将细胞核混悬液稀释于250 μl的IBM-Trion X-100中,轻加振荡混匀。为抗荧光褪色可加等量的抗褪色溶液至载片上的细胞核涂片上,选择适当的激发波长观察。(8)流式细胞计:将750 μl的细胞核混悬液通过流式细胞仪(Flow cytometry, FCM),DEMS处理过的红细胞作为对照(Df 530/30nm, Omega ·Optical Inc, Brattleboro, VT)。
荧光显微镜下H526细胞的慢病毒转染结果
慢病毒转染过后的细胞荧光照片明场[img]https://ng1.17img.cn/bbsfiles/images/2023/06/202306302120455921_65_5389809_3.jpeg[/img]低浓度[img]https://ng1.17img.cn/bbsfiles/images/2023/06/202306302120459963_6495_5389809_3.jpeg[/img]高浓度[img]https://ng1.17img.cn/bbsfiles/images/2023/06/202306302120461778_5983_5389809_3.jpeg[/img]
[font=宋体]哺乳动物细胞表达系统具有促使蛋白正确折叠和实现复杂修饰的功能,表达的蛋白更近天然状态,能够运用于制药等活性要求高的领域。利用哺乳动物细胞表达系统生产蛋白通常有两种方式:瞬时转染与稳定转染(构建稳定细胞系),这两种方式在原理、操作流程、应用场景上均有所区别,本文主要就瞬时转染与稳定转染之间的区别做一介绍。[/font][font=宋体] [/font][font=宋体][b]瞬时转染与稳定转染实验原理的异同:[/b][/font][font=宋体] [/font][font=宋体]瞬时转染与稳定转染都是将目的基因转染至特定哺乳动物细胞内,进而表达得到目的蛋白。不同的是瞬时转染的方式外源基因并没有转染到细胞的染色体上而是存在于游离的载体上,这样可以在短时间内获得基因的表达产物,但是随着细胞的不断分裂增殖外源基因最终会丢失,无法继续进行重组蛋白的生产;而利用细胞稳定转染则会将外源基因转染至细胞染色体上,目的基因不会随着细胞传代而消失,稳定转染的细胞株能够长期稳定的生产目的蛋白。[/font][font=宋体] [/font][font=宋体][b]实验操作的差别:[/b][/font][font=宋体][font=Calibri]1. [/font][font=宋体]瞬时转染与稳定转染所用的质粒是不同的,瞬转的质粒不需要带有抗性,而用于稳定转染的质粒一定要带有特定的抗性以便于后续的克隆株筛选。另外,两者所用的培养基及实验试剂也有所区别。[/font][/font][font=宋体][font=Calibri]2. [/font][font=宋体]瞬时转染的操作比构建稳定细胞系的操作简单,构建好质粒后,经过细胞复苏、转染、细胞培养、蛋白纯化等步骤即可得到目的蛋白;而构建稳定细胞系需要先将构建好的质粒线性化,再导入培养好的哺乳动物细胞内,通过一定的转染方式实现质粒与细胞的融合,接着经过细胞池筛选、单克隆筛选、细胞传代培养等步骤才能得到稳定转染的细胞系。对稳定细胞系进行培养能够长期稳定的生产目的蛋白。[/font][/font][font=宋体] [/font][font=宋体][b]瞬时转染与稳定转染优缺点对比[/b][/font][font=宋体] [/font][font=宋体]瞬时转染:[/font][font=宋体]优点:[/font][font=宋体]①能够快速生产得到微量至中量的重组蛋白[/font][font=宋体]②实验成本低[/font][font=宋体]③一个宿主可以带有多个拷贝,表达效率高[/font][font=宋体]缺点:无法长期生产得到重组蛋白[/font][font=宋体] [/font][font=宋体]稳定细胞系构建:[/font][font=宋体]优点:能够长期稳定生产目的蛋白[/font][font=宋体]得到稳转株之后后续生产蛋白的成本大大降低[/font][font=宋体]能够对基因进行基因插入、基因敲除等编辑操作[/font][font=宋体] [/font][font=宋体][font=宋体]义翘神州提供[url=https://cn.sinobiological.com/services/stable-cell-line-development-service][b]稳定细胞系构建服务[/b][/url],包含过表达细胞系构建服务和[/font][font=Calibri]CHO[/font][font=宋体]稳定细胞株开发服务,详情可以参看[/font][font=Calibri]https://cn.sinobiological.com/services/stable-cell-line-development-service[/font][/font]
常用的基因转染技术是将外源基因导入靶细胞需要一定的载体和导入方法,基因转技术则是将纯化的含有靶基因的质粒DNA送入细胞内,并在细胞内表达。转染方法有多种,根据不同的细胞,贴壁或悬浮细所可选用不同的方法,其目的是要达到设置转染效率,影响转染产率的因素有多种,包括转染方法、操作技术、质粒DNA的纯度、靶细胞的生长状态等,下面重点介绍向几种常用的转染技术:被用于作靶基因转染的细胞,其生长状态如何,将直接决定了基因转染效率。如为贴壁生长的细胞,一般要求在转染前一日,必需应用胰酶处理成单细胞悬液,重新接种于培养皿或瓶,细胞密度以铺满培养器皿的60%为宜,转染当日,在转染前4小时换一将近新鲜培养液。对于悬浮细胞,也需在转染前4小时换一次新鲜培养液。用于转染的质粒DNA必须无蛋白质,无RNA和其了化学物质的污染,OD260/280比值应在1.8以上。应用酚-氯仿抽提法制备的质粒DNA一般难以达到此标准,目前大多采用进口的术提取纯化试剂盒。具体的基因转染技术有鳞酸钙介导的转染法、DEAE葡聚糖介导转染法、脂质体介导转染法及电击基因转导尘等。靶基因被导入细胞后,一般在转染后48小时,靶基因即在细胞内表达。根据不同的实验目的,48小时后即可进行靶基因表达的检测等实验。如若建立稳定的细胞系,则可对靶细胞进行筛选,根据不同基因载体中所含有的抗性标志选用相应的药物,最常用的直核表达基因载体的标志物有潮霉素(hygromycin)和新霉素(neomycin)。
1、 电场参数电场是电转染的重要因素,细胞在电场的作用下,膜通透性增加或是形成小孔,以完成转染过程。因此电场强度是应该被优化的主要参数。电场强度不能过高,过高会增加细胞的死亡率;也不能过低,过低不能增加膜的通透性或在膜上形成小孔。因此,一个适宜的电场强度至关重要。不同细胞系具有不同的最佳场强值,其确定方法除了实验直接测定比较不同场强下转染率的高低外,还可以采取较为简便的间接法。文献显示存活率在50%左右的电场参数为理想参数,故可间接测定存活率来确定最佳场强值。
[font=宋体]随着生物技术的快速发展,稳定转染细胞株的构建在基础研究和应用研究中变得越来越重要。这一技术涉及到将外源基因稳定整合到宿主细胞基因组中,从而实现基因的长期表达。然而,这一过程也伴随着一系列的挑战和常见问题。本文将深入探讨稳转细胞株构建的方法,以及在实践过程中可能遇到的问题,旨在为研究者提供实用的指导和建议。[/font][font=宋体] [/font][font=宋体][b]稳定转染细胞株构建方法:[/b][/font][font=宋体] [/font][font=宋体]一、慢病毒感染细胞[/font][font=宋体] [/font][font=宋体][font=宋体]慢病毒因为可以感染大多数的分裂细胞和非分裂细胞,包装技术成熟,[/font][font=Calibri]II[/font][font=宋体]代和[/font][font=Calibri]III[/font][font=宋体]代慢病毒包装系统均能包装高滴度的慢病毒,是稳转细胞系构建的首选系统。但是因为慢病毒有包装容量限制,不适合转录本区域比较长的基因,对较大的基因有局限性。另外由于慢病毒是逆转录病毒,病毒表达载体中,是由[/font][font=Calibri]WPRE[/font][font=宋体]元件代替[/font][font=Calibri]PolyA[/font][font=宋体]稳定以达到稳定[/font][font=Calibri]mRNA[/font][font=宋体]的作用,对部分基因的翻译有一定影响。[/font][/font][font=宋体] [/font][font=宋体]二、转座子介导的稳转细胞系构建[/font][font=宋体] [/font][font=宋体][font=宋体]转座子是存在于各种生命细胞内的可移动遗传信息元件,能实现[/font][font=Calibri]DNA[/font][font=宋体]片段在染色体内部[/font][font=Calibri]/[/font][font=宋体]之间的转移。工程化的转座子由[/font][font=Calibri]ITR[/font][font=宋体]和转座酶编码基因组成,通过转座酶与[/font][font=Calibri]ITR[/font][font=宋体]结合实现基因转移。将转座子的两个元件分别构建在两个独立载体上,并将目的基因序列替换转座酶序列,与另一个载体共转染目的细胞,实现目的基因在宿主基因组上的整合。与其他技术不同,目的基因在整合酶存在下会持续处于切割—整合状态,实现连续跳跃。[/font][/font][font=宋体] [/font][font=宋体] [/font][font=宋体][font=宋体]三、[/font][font=Calibri]CRISPR/Cas9 [/font][font=宋体]基因敲入[/font][/font][font=宋体] [/font][font=宋体][font=宋体]利用[/font][font=Calibri]CRISPR/Cas9[/font][font=宋体]系统,通过[/font][font=Calibri]NHEJ[/font][font=宋体]和[/font][font=Calibri]HDR[/font][font=宋体]修复机制,可将目的基因(如抗性基因和荧光标志)定点敲入细胞基因组。在哺乳动物细胞中,[/font][font=Calibri]NHEJ[/font][font=宋体]的效率高于[/font][font=Calibri]HDR[/font][font=宋体]。设计特定[/font][font=Calibri]sgRNA[/font][font=宋体]和[/font][font=Calibri]donor[/font][font=宋体]载体,可在[/font][font=Calibri]AAVS1[/font][font=宋体]位点(人基因组安全港)实现基因定点敲入。此方法稳定、安全,但敲入概率在不同细胞和位点中有所差异,需筛选单克隆以获得稳定细胞系。随着编辑效率提高,此方法将更省时省力。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]四、[/font][font=Calibri]CRISPR SAM[/font][font=宋体]系统,通过融合[/font][font=Calibri]VP64[/font][font=宋体]、[/font][font=Calibri]MS2-P65-HSF1[/font][font=宋体]系统对基因进行过表达和干扰[/font][/font][font=宋体][font=Calibri]CRISPR SAM[/font][font=宋体]通过[/font][font=Calibri]dCas9-VP64[/font][font=宋体]、[/font][font=Calibri]MS2-P65-HSF1[/font][font=宋体]激活蛋白实现了多数细胞内源基因的特异性激活,不受基因大小限制。主要优势是应用广泛,但需要三种不同抗性病毒逐次或同时感染细胞。[/font][font=Calibri]CRISPR SAM[/font][font=宋体]将[/font][font=Calibri]sgRNA[/font][font=宋体]序列构建在[/font][font=Calibri]lenti[/font][font=宋体]载体中,配合其他两种慢病毒可激活任何基因。主要缺点是筛选细胞时需要三种不同抗性的病毒,部分细胞筛选后表型变化明显。[/font][/font][font=宋体] [/font][font=宋体] [/font][font=宋体][b]稳转细胞系构建中常见问题解析:[/b][/font][font=宋体] [/font][font=宋体][font=Calibri]1. [/font][font=宋体]是否需要进行密码子优化?[/font][/font][font=宋体] [/font][font=宋体][font=宋体]由于氨基酸密码子的简并性和[/font][font=Calibri]tRNA[/font][font=宋体]的种类多样性、数量的差异,密码子优化对长基因稳转细胞系的构建有很强的必要性,可以大幅提高目标蛋白的表达量。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]2. Kozak[/font][font=宋体]序列是必须要添加的吗?[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]Kozak[/font][font=宋体]序列(通常是[/font][font=Calibri]GCCACCatgG[/font][font=宋体]),真核生物[/font][font=Calibri]mRNA[/font][font=宋体]真正的起始密码子位于[/font][font=Calibri]Kozak[/font][font=宋体]序列的保守序列中,其共有序列是[/font][font=Calibri]CCRCCAUGG[/font][font=宋体],几乎所有[/font][font=Calibri]mRNA[/font][font=宋体]的翻译起始都依赖于[/font][font=Calibri]5[/font][font=宋体]’帽子结构募集小亚基,少数通过内部核糖体进入位点[/font][font=Calibri](IRES)[/font][font=宋体]募集小亚基。[/font][font=Calibri]Kozak[/font][font=宋体]序列通过与翻译起始因子[/font][font=Calibri]eIF[/font][font=宋体]形成翻译起始复合物,起始蛋白的翻译。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]3. [/font][font=宋体]启动子应如何选择?[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]CMV[/font][font=宋体](巨细胞病毒)启动子除了在干细胞外,其他大多数细胞类型中均可以获得高表达活性。悬浮细胞中[/font][font=Calibri]SFFV[/font][font=宋体]启动子表达活性相对较高。而[/font][font=Calibri]EF1a[/font][font=宋体](延伸因子[/font][font=Calibri]-1[/font][font=宋体]α)启动子更适用于表达长片段的基因。[/font][font=Calibri]CMV[/font][font=宋体];[/font][font=Calibri]EF1A[/font][font=宋体];[/font][font=Calibri]CAG[/font][font=宋体];[/font][font=Calibri]CBh[/font][font=宋体];[/font][font=Calibri]SFFV[/font][font=宋体],等均属于强启动子,都可作为稳转细胞系构建可选启动子。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]4. [/font][font=宋体]稳转细胞系培养体系[/font][/font][font=宋体] [/font][font=宋体][font=宋体]稳转细胞系一般建议用半抗性筛选浓度维持细胞生长,如[/font][font=Calibri]hepG2[/font][font=宋体]细胞[/font][font=Calibri]puro[/font][font=宋体]抗性筛选浓度为[/font][font=Calibri]2[/font][font=宋体]μ[/font][font=Calibri]g/ml[/font][font=宋体],则建议用[/font][font=Calibri]1ug/mL+[/font][font=宋体]完全培养基维持生长;部分基因过表达后会影响细胞代谢,尤其肿瘤细胞,糖酵解代谢速率受影响,细胞生长缓慢,可相应添加葡萄糖或者[/font][font=Calibri]HEPES[/font][font=宋体]调节细胞细胞代谢水平,维持细胞增殖。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]5. [/font][font=宋体]标签放在[/font][font=Calibri]N[/font][font=宋体]端还是[/font][font=Calibri]C[/font][font=宋体]端好?[/font][/font][font=宋体] [/font][font=宋体][font=宋体]如果[/font][font=Calibri]N[/font][font=宋体]端有信号肽,建议放在[/font][font=Calibri]C[/font][font=宋体]端;蛋白如果较小,建议放在[/font][font=Calibri]C[/font][font=宋体]端,减少蛋白降解;如果下游有[/font][font=Calibri]P2A[/font][font=宋体],[/font][font=Calibri]T2A[/font][font=宋体]等自剪切肽,建议放在[/font][font=Calibri]C[/font][font=宋体]端。如果为了添加[/font][font=Calibri]Kozak[/font][font=宋体]序列,建议加在[/font][font=Calibri]N[/font][font=宋体]端。如果纯化中需要酶切标签,建议放在[/font][font=Calibri]N[/font][font=宋体]端。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]6. [/font][font=宋体]是否可以构建稳转敲除的细胞系?[/font][/font][font=宋体] [/font][font=宋体]一般不建议,因为敲除元件稳定持续的表达在细胞中,累积脱靶效应明显,影响细胞状态。[/font][font=宋体] [/font][font=宋体][font=宋体]义翘神州提供[url=https://cn.sinobiological.com/services/stable-cell-line-development-service][b]稳定细胞系构建服务[/b][/url],包含过表达细胞系构建服务和[/font][font=Calibri]CHO[/font][font=宋体]稳定细胞株开发服务,其服务内容详情可以参看:[/font][font=Calibri]https://cn.sinobiological.com/services/stable-cell-line-development-service[/font][/font]
[color=#1a1a1a]1. 脂质体法[/color][color=#1a1a1a][color=#1a1a1a]2. 电穿孔法[/color][/color][color=#1a1a1a][color=#1a1a1a][color=#1a1a1a]3. 病毒介导的感染[/color][/color][/color][color=#1a1a1a][color=#1a1a1a][color=#1a1a1a]4.非脂质体转染[/color][/color][/color][color=#1a1a1a][color=#1a1a1a][color=#1a1a1a].........请大家补充[/color][/color][/color]
4、 质粒因素从质粒浓度看,细胞密度为1*106/ml,DNA用量在2-5ug/ml转染效率最高。转染率在一定一定范围内随质粒浓度的上升呈线性增高,但是到达一个峰值后,随DNA用量增加而转染率逐渐下降,其原因可能为细胞吸纳DNA有一个饱和度,过量便会产生毒性,使存活率降低,不利于转染率提高。进行小分子量的分子转染时(siRNA/miRNA),应使用高电压、短脉冲时间。大分子量分子转染时(DNA),应使用低电压,长脉冲时间。从质粒的纯度看,用高纯度的DNA才 能提高电转的效率,且应注意以下几点:首先DNA/RNA应该超纯(A260/A2801.8),其次应不含内毒 素,同时质粒应溶于双蒸水而不是TE缓冲溶液。
[b][url=http://www.f-lab.cn/cell-analyzers/puncher.html][b]单细胞转移分离系统[/b][/url]是可用于单细胞转移,单细胞分离和单细胞隔离,单细胞成像应用的多功能单细胞分离操作仪器,它可以实现从微孔芯片转移单细胞到细胞收集管中。单细胞转移分离系统[/b][color=#666666]集单细胞成像,单细胞隔离,单细胞选择功能于一体,自动聚焦成像。[/color][b]单细胞转移分离系统转移单细胞到Eppendorf微管,PCR微孔板或其它反应微管中,[/b][color=#666666]在隔离单细胞后,它可以对选定收集的细胞进行扫描并成像。[/color][b]单细胞转移分离系统[/b][color=#666666]采用Nikon Ti-2倒置荧光显微镜,配备自动扫描显微镜载物台,自动聚焦器件,高灵敏度荧光CCD相机和LED激发光源组建而成。[/color][img=单细胞转移分离系统]http://www.f-lab.cn/Upload/single-cell-isolation.JPG[/img][b]单细胞转移分离系统[/b]特点完全自动化,步进系统高质量单细胞荧光成像单细胞分离的效率超过90% 超过70%分离的细胞增殖 分离后兼容所有的单细胞的WGA工具包(放大器的‐1,picoplex,复制‐G)实惠微Wells基于硅微孔微腔。由薄膜封闭70µ m,井底直径(1µ m),包含一个单孔。样品流体进入威尔斯并从底部的孔隙中流出。单个细胞被拖着走。一旦单个细胞降落到孔隙上,流动停止,其他细胞就不会进入井内。有用的细胞被识别出来。选定的细胞穿孔从微孔到384孔PCR板或离心管等等。单细胞转移分离系统:[url]http://www.f-lab.cn/cell-analyzers/puncher.html[/url]
6、电转染试剂的选择电击会对细胞造成一定程度的伤害,在转染试剂的选择上要注意,应选择具有细胞膜修复成分的电转染试剂,如Entranster-E,将电击对细胞的损伤降到最低。
5、 温度 一般情况下,电转的过程是在室温下进行,但在 电击前后可对细胞进行冰浴处理。电击前冰浴的时间对转染率影响不大,但低温环境能够防止DNA被外源性DNA酶降解,并限制在电击中因Joule作用而产生的不良反应,因此,实验一般选择电击前冰浴5min。电击后冰浴,会影响DNA摄入,因为电击后冰浴可增强细胞收缩,细胞膜上的 电穿孔封闭较慢,延长外源性DNA摄入时间,从而提高其摄入量。在室温环境下,虽然细胞膜上的孔 封闭速度快,但在随后2h,质粒还可通过电内化进入细胞,因此摄人量不一定比4℃环境下低。而且,室温下细胞在5 min内即闭孔,在4℃的环境下穿孔状态可保持4h,穿孔封闭缓慢降低了存活率,同时细胞在低温下更容易受到损伤。因此,综合考虑摄人量和存活率,大部分文献倾向于电击后细胞仍在室温或37℃下保存。
哪位大神做过生物方面,如细胞的核磁谱,可否请教下经验,跟普通样品有什么不一样,需要注意什么?
2、脉冲过程1)脉冲波形脉冲波形主要分为两种:1、方波脉冲;2、指数递减波脉冲。方波脉冲是指:电压瞬间升至预设电压,保持电压放电,然后瞬间终止放电。一般哺乳动物细胞电转染时选择方波脉冲,有较高的转染效率和细胞存活率。指数递减波脉冲是指:先对电容充电,然后让电容完全放电,其电压变化呈指数递减。一般这种波形的脉冲电转染适用于细菌、酵母菌、昆虫细胞。2)脉冲时间脉冲时间的选定主要取决与脉冲波形。在方波脉冲中,脉冲时间可直接设定。在指数递减波脉冲中,脉冲时间是指电压衰减至初始电压1/3时所用的时间,等于电容(C)与电阻(R)的乘积,单位是ms。在参数优化中,增加电压应当降低脉冲时间,而减小电压则应当增大脉冲时间。3)脉冲次数一般而言,对于大多数细胞类型都选择单次脉冲。而在有些情况下可能会用到多次脉冲,因为低电压、短脉冲时间、多次脉冲可有效避免细胞损伤。多次脉冲建议中间间隔1min。
http://gene.bjmu.edu.cn/news/ap1.gif 细胞凋亡与坏死是两种完全不同的细胞凋亡形式,根据死亡细胞在形态学、生物化学和分子生物学上的差别,可以将二者区别开来。细胞凋亡的检测方法有很多,下面介绍几种常用的测定方法。 一、细胞凋亡的形态学检测 根据凋亡细胞固有的形态特征,人们已经设计了许多不同的细胞凋亡形态学检测方法。 1 光学显微镜和倒置显微镜 (1) 未染色细胞:凋亡细胞的体积变小、变形,细胞膜完整但出现发泡现象,细胞凋亡晚期可见凋亡小体。 贴壁细胞出现皱缩、变圆、脱落。 (2) 染色细胞:常用姬姆萨染色、瑞氏染色等。凋亡细胞的染色质浓缩、边缘化,核膜裂解、染色质分割 成块状和凋亡小体等典型的凋亡形态。 2 荧光显微镜和共聚焦激光扫描显微镜 一般以细胞核染色质的形态学改变为指标来评判细胞凋亡的进展情况。 常用的DNA特异性染料有:HO 33342 (Hoechst 33342),HO 33258 (Hoechst 33258), DAPI。三种染料与 DNA的结合是非嵌入式的,主要结合在DNA的A-T碱基区。紫外光激发时发射明亮的蓝色荧光。 Hoechst是与DNA特异结合的活性染料,储存液用蒸馏水配成1mg/ml的浓度,使用时用PBS稀释成终浓度为2~5mg/ml。 DAPI为半通透性,用于常规固定细胞的染色。储存液用蒸馏水配成1mg/ml的浓度,使用终浓度一般为0.5 ~1mg/ml。 结果评判:细胞凋亡过程中细胞核染色质的形态学改变分为三期:Ⅰ期的细胞核呈波纹状(rippled)或呈折缝样(creased),部分染色质出现浓缩状态;Ⅱa期细胞核的染色质高度凝聚、边缘化;Ⅱb期的细胞核裂解为碎块,产生凋亡小体(图1)。 3 透射电子显微镜观察 结果评判:凋亡细胞体积变小,细胞质浓缩。凋亡Ⅰ期(pro-apoptosis nuclei)的细胞核内染色质高度盘绕,出现许多称为气穴现象(cavitations)的空泡结构(图2);Ⅱa期细胞核的染色质高度凝聚、边缘化;细胞凋亡的晚期,细胞核裂解为碎块,产生凋亡小体。
摘要:细胞凋亡与坏死是两种完全不同的细胞凋亡形式,根据死亡细胞在形态学、生物化学和分子生物学上的差别,可以将二者区别开来。细胞凋亡的检测方法有很多,下面介绍几种常用的测定方法。一、细胞凋亡的形态学检测根据凋亡细胞固有的形态特征,人们已经设计了许多不同的细胞凋亡形态学检测方法。1. 光学显微镜和倒置显微镜(1) 未染色细胞:凋亡细胞的体积变小、变形,细胞膜完整但出现发泡现象,细胞凋亡晚期可见凋亡小体。贴壁细胞出现皱缩、变圆、脱落。(2) 染色细胞:常用姬姆萨染色、瑞氏染色等。凋亡细胞的染色质浓缩、边缘化,核膜裂解、染色质分割成块状和凋亡小体等典型的凋亡形态。2. 荧光显微镜和共聚焦激光扫描显微镜一般以细胞核染色质的形态学改变为指标来评判细胞凋亡的进展情况。常用的DNA特异性染料有:HO 33342 (Hoechst 33342),HO 33258 (Hoechst 33258), DAPI。三种染料与 DNA的结合是非嵌入式的,主要结合在DNA的A-T碱基区。紫外光激发时发射明亮的蓝色荧光。Hoechst是与DNA特异结合的活性染料,储存液用蒸馏水配成1mg/ml的浓度,使用时用PBS稀释成终浓度为2~5mg/ml。DAPI为半通透性,用于常规固定细胞的染色。储存液用蒸馏水配成1mg/ml的浓度,使用终浓度一般为0.5 ~1mg/ml。结果评判:细胞凋亡过程中细胞核染色质的形态学改变分为三期:Ⅰ期的细胞核呈波纹状(rippled)或呈折缝样(creased),部分染色质出现浓缩状态;Ⅱa期细胞核的染色质高度凝聚、边缘化;Ⅱb期的细胞核裂解为碎块,产生凋亡小体(图1)。 http://img.dxycdn.com/trademd/upload/userfiles/image/2013/08/A1376376278_small.jpg3 透射电子显微镜观察结果评判:凋亡细胞体积变小,细胞质浓缩。凋亡Ⅰ期(pro-apoptosis nuclei)的细胞核内染色质高度盘绕,出现许多称为气穴现象(cavitations)的空泡结构(图2);Ⅱa期细胞核的染色质高度凝聚、边缘化;细胞凋亡的晚期,细胞核裂解为碎块,产生凋亡小体。 http://img.dxycdn.com/trademd/upload/userfiles/image/2013/08/A1376376295_small.jpg二、磷脂酰丝氨酸外翻分析(Annexin V法)磷脂酰丝氨酸(Phosphatidylserine, PS)正常位于细胞膜的内侧,但在细胞凋亡的早期,PS可从细胞膜的内侧翻转到细胞膜的表面,暴露在细胞外环境中(图3)。Annexin-V是一种分子量为35~36KD的Ca2+依赖性磷脂结合蛋白,能与PS高亲和力特异性结合。将Annexin-V进行荧光素(FITC、PE)或biotin标记,以标记了的Annexin-V作为荧光探针,利用流式细胞仪或荧光显微镜可检测细胞凋亡的发生。碘化丙啶(propidine iodide, PI)是一种核酸染料,它不能透过完整的细胞膜,但在凋亡中晚期的细胞和死细胞,PI能够透过细胞膜而使细胞核红染。因此将Annexin-V与PI匹配使用,就可以将凋亡早晚期的细胞以及死细胞区分开来。http://img.dxycdn.com/trademd/upload/userfiles/image/2013/08/A1376376296_small.jpg方法1. 悬浮细胞的染色:将正常培养和诱导凋亡的悬浮细胞(0.5~1×106)用PBS洗2次,加入100 ul Binding Buffer和FITC标记的Annexin-V(20ug/ml)10 ul,室温避光30 min,再加入PI(50 ug/ml)5 ul,避光反应5 min后,加入400 ul Binding Buffer,立即用FACScan进行流式细胞术定量检测(一般不超过1 h), 同时以不加AnnexinV-FITC及PI的一管作为阴性对照。2. 贴壁培养的细胞染色:先用0.25%的胰酶消化,洗涤、染色和分析同悬浮细胞。3. 爬片细胞染色:同上,最后用荧光显微镜和共聚焦激光扫描显微镜进行观察。结果 http://img.dxycdn.com/trademd/upload/userfiles/image/2013/08/A1376376298_small.jpg http://img.dxycdn.com/trademd/upload/userfiles/image/2013/08/A1376376300_small.jpg
3、细胞因素用于电转的细胞一般选取处于对数生长期的细胞(15代以内,传代后2d)。因为处于对数生长期的细胞分裂旺盛,表面结构致密比稳定期的细胞差,电转后,细胞膜的恢复能力强,而且处于有丝分裂期的细胞更容易接受外源DNA.细胞悬液浓度一般为1*106/ml,当细胞生长密度大于3*106/ml,转染效率会下降。原因除了细胞老化外,细胞过密会使相邻细胞相互作用增大,甚至使细胞相互融合,从而导致电场的局部微扰,使电场环境无法均一化,细胞体积越大对电击越敏感,所需电场强度也越小。
900万像素显微镜摄像头加在生物显微镜上所拍细胞核移植显微图片:[IMG]http://mshot.cn//uploadfile/b/RxCGPWaeCYhTU1tWiNYo.jpg[/IMG]
[b][font=宋体]前言[/font][/b][font=宋体]在蛋白质研究领域,稳定细胞系的应用已成为生产高质量结构生物学蛋白质的关键手段。随着技术的不断进步,稳定细胞系的生成与筛选方法得到了显著改进,从而推动了蛋白质生产的高效化与精准化。[/font][font=Calibri] [/font][b][font=宋体]细胞系的建立和应用[/font][font=宋体][font=Calibri]HEK293[/font][font=宋体]和[/font][font=Calibri]CHO[/font][font=宋体]细胞系[/font][/font][/b][font=宋体]因其稳定的蛋白表达和适当的翻译后修饰而被广泛用于结构生物学研究。这些细胞系能有效地生产具有复杂糖基化模式的蛋白质,这对于确保蛋白质的功能和稳定性至关重要。糖基化缺陷细胞系通过特定的基因改造,能够分泌脱糖基化糖蛋白,为蛋白质生产提供了更加纯净的原料。[/font][font=Calibri] [/font][b][font=宋体]稳定细胞系的生成[/font][/b][font=宋体][font=宋体]传统的稳定细胞系生成技术如瞬时转染,虽然方法简便,但存在整合频率低、转基因沉默等问题。为了克服这些困难,研究者们开发出了一系列新技术,如细胞分选技术、位点特异性重组(如[/font][font=Calibri]FLP/FRT[/font][font=宋体]系统)、转座子系统(如[/font][font=Calibri]piggyBac[/font][font=宋体])、慢病毒系统以及噬菌体整合酶等,提高了稳定细胞系的生成效率和稳定性。[/font][/font][font=宋体] [/font][font=宋体][font=宋体]序列特异性基因组工程也为稳定细胞系的生成提供了新的思路。通过敲除或修饰特定的基因,研究者们能够实现对细胞功能的精准调控,从而优化蛋白质生产的效率和纯度。例如,一种同时缺乏[/font][font=Calibri]GnTI[/font][font=宋体]和谷氨酰胺合成酶([/font][font=Calibri]GS[/font][font=宋体])活性的[/font][font=Calibri]CHO[/font][font=宋体]细胞系被成功开发出来,为高效筛选具有[/font][font=Calibri]GS[/font][font=宋体]标记的稳定细胞系提供了有力工具。[/font][/font][font=Calibri] [/font][b][font=宋体]稳定细胞系与瞬时转染的比较[/font][/b][font=宋体]稳定细胞系相较于瞬时转染具有多个优点,包括能够进行大规模生产和保持高水平的蛋白表达稳定性。尽管瞬时转染在某些情况下能快速产生大量蛋白,但其表达水平和重复性通常不如稳定细胞系。[/font][font=Calibri] [/font][b][font=宋体]展望[/font][/b][font=宋体]近年来,利用稳定细胞系高效生产结构生物学蛋白质已成为研究的热点和趋势。通过引入新技术、优化筛选方法和改进整合系统,不仅能够提高蛋白质生产的效率和纯度,还能够为结构生物学研究提供更加精准、可靠的实验工具。随着基因编辑和细胞工程技术的进步,预计在未来,通过精确的基因操作能够更有效地创建和利用稳定细胞系。这些技术的进步将促进结构生物学和药物开发中蛋白质的高效和可持续生产。[/font][font=宋体] [/font][font=宋体]本文由义翘神州进行整理,同时提供[/font][url=https://cn.sinobiological.com/services/stable-cell-line-development-service][u][font=宋体][color=#0000ff]稳定细胞系构建服务[/color][/font][/u][/url][font=宋体],详情可点击了解![/font][font=Calibri] [/font][font=宋体]参考文献:[/font][font=Calibri]Büssow K. Stable mammalian producer cell lines for structural biology. [/font][i][font=Calibri]Curr Opin Struct Biol[/font][/i][font=Calibri]. 2015 32:81-90. doi:10.1016/j.sbi.2015.03.002[/font]
iPSC神经细胞培养试剂【胜创生物】成功取得GlobalStem中国区总代理:人ES细胞mRNA高效转染试剂,GlobalStem公司2006年在美国成立,其团队核心技术骨干为原Lifetech部门,负责研发生产细胞转染试剂Lipofectamine 2000、Lipofectamine 3000及细胞培养试剂等。 GlobalStem专注于iPSC(诱导性多功能干细胞神经分化)神经细胞转染、干细胞转染试剂,iPSC神经细胞培养、干细胞培养、原代细胞培养试剂。 【胜创生物】http://www.shengchuangbio.com/-人ES细胞mRNA高效转染试剂 DNA-In-Neuro:在神经细胞中的转染效率约高于Lipofectamine 2000的 3倍。 DNA-In-Neuro:*转染效率高*低毒性*数据重复性高*操作简单。 日前,【胜创生物】公司已成功取得GlobalStem的中国总代理权,电话咨询400-6400-850 胜创生物以“品质第一,客户第一“为公司价值观,以引进”新产品、新技术“为己任,更多资讯关注胜创生物微信【sc-bios】获取详情!
摘要激光扫描共聚焦显微镜作为80年代发展起来的一种高精度分子细胞生物学分析仪器,具有组织细胞断层扫描、活细胞动态荧光监测、三维图像重建、共聚焦图像定量分析等先进功能,在近年的细胞凋亡这一研究热点中得到了大量创造性的应用。本文拟就对激光扫描共聚焦显微镜在凋亡的形态学、分子水平变化及重要生理过程三方面研究中的应用及其成果做一综述。细胞凋亡(apoptosis)是不同于细胞坏死的一种细胞主动死亡方式,并由特定的基因控制。凋亡细胞在形态上出现变圆皱缩、染色质浓缩边集、核碎裂、凋亡小体形成等变化,并最终由非炎症过程清除。由于细胞凋亡独特地影响着机体的细胞发育和代谢,在监测和清除肿瘤细胞与突变细胞等方面也可能发挥重要的作用,近年来受到了细胞生物学、分子生物学、免疫学等多学科的广泛关注。激光扫描共聚焦显微镜(laser scaing confocal microscopy, LSCM)是80年代发展起来的一种高精度分子细胞生物学分析仪器,辅以各类免疫荧光探针或荧光染料与被测物质特异性结合,不仅可观察固定的细胞组织切片,还可对活细胞的结构、分子和离子进行实时动态地观察和检测。在细胞凋亡的研究中,激光扫描共聚焦显微镜已被广泛地应用于形态学、分子水平监测及重要生理改变等各方面,其中不乏新颖之处,并获得了大量成果,以下将就此做一简单的介绍。激光扫描共聚焦显微镜与凋亡的形态学激光扫描共聚焦显微镜用点光源扫描标本的光学横断面,以代替普通光学显微镜所使用的场光源,并用探测针孔滤去离焦光线,所以消除了来自焦平面以外的衍射或散射光的干扰,可实现高清晰、高分辨率的组织细胞断层扫描。并且由于激光扫描共聚焦显微镜采用数字化成像,因而辅以一定的软件就能对图像进行定量分析及三维重建等操作。过去对细胞凋亡的形态学研究方法局限于活性细胞和组织切片染色、荧光镜观察,或者石蜡切片原位末端标记法。由于普通光镜的分辨率和清晰度有限,而电镜又显然不适合对凋亡这一复杂动态过程的监测,激光扫描共聚焦显微镜的应用使人们对细胞凋亡的形态学观察分析提高到了一个前所未有的新水平。细胞核核膜的破坏对于染色质聚集并形成凋亡小体起重要作用。lamin是构成核片层的蛋白质,位于核膜的内表面,由caase-6介导的lamin裂解可影响核膜的完整性。在McCall等的研究中,对果蝇卵子发生晚期的细胞凋亡现象进行了动态观察。以单抗mAb101标记其哺育细胞核内膜的laminDm0(哺乳类laminB的同源体),用激光扫描共聚焦显微镜加以观察。正常哺育细胞到11期时,染色的lamin呈弥散的雾状分布并围绕核周,而dcp-1GLC哺育细胞即使到了较晚的14期时,仍然显示界线明确的染色。可见dcp-1突变体在核lamin蛋白的酶切或解聚方面存在缺陷。细胞器Li 等在对C(6)-酰基鞘氨醇诱导胞内囊泡产生的研究中,在不产生中毒效应的情况下,加入10microM C(6)-酰基鞘氨醇以诱导鼠纤维母细胞(3T3-L1和3T3-F442A)凋亡。观察到囊泡的形成与C(6)-酰基鞘氨醇的诱导呈时间依从和剂量依从关系。大量小泡在其加入后8小时内出现,并且随时间而增大;大泡最终分布在核周,而小泡分布在细胞边缘。用抗-溶酶体膜蛋白抗体和共聚焦免疫荧光显微分析,证明增大的囊泡为晚期内吞体/溶酶体。另外,胞内的细胞器都有其适用的荧光探针,如高尔基复合体常用的探针有Dceramide、BODIPY ceramide等,内质网常用的有Dil、DiOC6等,经标记均可进行精细的观察。当然,激光扫描共聚焦显微镜在形态学中的优势更在于其对图像的三维重建功能,从而揭示过去只能在平面上显现的凋亡细胞在三维空间中的结构;而对细胞凋亡的动态过程,它可以用三维加时间的四维方式进行观察,来获取最逼真的形态学资料。凋亡过程中一些特征性的三维形态变化正期待着进一步具体的工作去发现。激光扫描共聚焦显微镜对凋亡细胞的分子水平监测随着分子生物学突飞猛进的发展,关于细胞凋亡分子机制的研究已有了很大的突破。细胞凋亡的信号传递途径及其调控涉及到大量的酶级联反应、生物大分子的空间转移等。而激光扫描共聚焦显微镜以其定性、定量、定时的优点,结合众多荧光探针的应用,成为了研究细胞凋亡分子水平变化的有力手段。DNA大分子DNA断裂以及染色质的异常凝聚,是细胞凋亡的关键,同时也是细胞核在细胞凋亡中具有标志性的变化。Columbara等报道将激光扫描共聚焦显微镜与原位TdT和Poll免疫荧光技术相结合,进而确定双链和单链DNA的断裂点。而在对细胞凋亡和细胞坏死区别的研究中,Kreel等在培养的K562细胞中加入放线菌素D以诱导凋亡,并对细胞的DNA片段进行了3’-末端标记。经激光扫描共聚焦显微镜观察发现K562细胞凋亡早期有大量DNA片段出现,且DNA片段弥散分布于除核仁外的细胞核区。伴随着凋亡的进展,细胞核内出现大量高标记密度的圆形小体。而采用NaN3或快速冻融法使细胞坏死,经激光扫描共聚焦显微镜观察证实,在坏死开始阶段并无DNA片段的出现,至少在坏死发生24小时后才有DNA片段产生。Caase家族Caases是一组高度保守的半胱氨酸蛋白酶,目前发现有11个成员。多数细胞凋亡是以Caase家族蛋白的激活并作用于其关键底物而实现的,而caases激活的关键又在于该家族蛋白间的级联反应,因此caases被认为是细胞凋亡的中心环节和执行者,成为研究的热点。Mandal等用激光扫描共聚焦显微镜对细胞凋亡中激活的caase-3的重分布进行了研究。用丁酸处理细胞后,观察到DNA-PKcs的裂解与caase-3的激活成正相关,而Bcl-2的过度表达则可抑制上述两个过程。同时还证明(1)激活后的caase-3重分布到核区,(2)裂解局部的DNA-PKcs和PARP(polyADP-ribosepolymerase,聚腺苷二磷酸核糖多聚酶),(3)裂解产物又被释放到核外的细胞液。caase-3的抑制物四肽DEVD-CHO又可抑制上述的三个连续的步骤。该研究提示:激活的caase-3在核内的重分布构成了丁酸所诱导的细胞凋亡中的一个重要凋亡信号。另外,在用激光扫描共聚焦显微镜对Q79诱导大鼠神经元凋亡的研究中,Sanchez等发现了Q79对caase-8的聚集和激活,而对caase-8的抑制则阻止了被诱导的细胞凋亡;加以Westernblot分析,还建立了caase-8的激活和某些神经退行性疾病(如舞蹈病)的联系。Grazyme丝氨酸蛋白酶grazyme为另一种重要的凋亡信号分子,对某些caase家族蛋白也有激活作用。Trapani等就证明了杀伤淋巴细胞利用穿孔素和grazymeB的协同作用来诱导靶细胞的凋亡,在其研究中通过激光扫描共聚焦显微镜观察到(1)50%细胞的胞核内快速聚集了以FITC荧光标记的grazymeB(最长7分钟,t1/2为2分钟),然后发生凋亡;(2)其它的细胞只有细胞液内有FITC-grazyme B的摄取,避免了凋亡。此间至少在13分钟后才有DNA碎片的出现,说明核内的grazyme B聚集出现在凋亡的执行阶段之前。并且通过对核内液的处理(加入70KDa FITC-dextran),间接观察到grazyme B的转移并非是因为核膜受caases的作用而破损,而是由于穿孔素的协同。其它以上的介绍显示,激光扫描共聚焦显微镜在检测活细胞酶活性动态变化方面有着突出的优势。实际上,对于细胞凋亡的分子机制这样一个极其复杂的课题,激光扫描共聚焦显微镜的应用远不只限于上述的几种离子和大分子,而是渗透到了大量的分枝课题中去。如在对重要的凋亡负调控蛋白Bcl-2的研究中,Beham等利用基因毒性损害(genotoxic damage)诱导细胞凋亡,并以Bcl-2蛋白抑制其凋亡过程。用激光扫描共聚焦显微镜和Immunoblotting观察显示,Bcl-2的作用在于阻止了诱导产生的p53蛋白向核内的转运。而Ohsawa等对独立于caase家族的另一种重要蛋白酶—组织蛋白酶进行了研究,用血清剥夺法诱导PC12细胞凋亡,并用激光扫描共聚焦显微镜监测了其精细超微结构改变过程和细胞内组织蛋白酶B和D的免疫活度的对比变化。又如,在人胰岛淀粉样多肽(hIA)的研究中,Hiddinga等用表达hIA的质粒转染COS-1细胞诱导凋亡,辅以免疫组化染色,用激光扫描共聚焦显微镜证明了hIA在细胞的内质网和高尔基复合体内呈簇状沉积,并与细胞
[font=宋体]慢病毒构建稳转细胞系的原理主要是利用慢病毒载体将外源基因导入宿主细胞,并实现外源基因的稳定表达。具体来说,构建稳转细胞系的核心是将慢病毒矢量载体导入宿主细胞中,慢病毒载体通常包含病毒的复制和包装组件,以及外源基因的表达调控序列。当慢病毒载体被导入宿主细胞后,它可以利用细胞的复制和转录机制将外源基因插入宿主细胞的染色体中,从而实现外源基因的稳定表达。[/font][font=宋体] [/font][font=宋体][b]构建稳定的慢病毒转染细胞系是在细胞中稳定表达外源基因的一种有效方法。下面是一般慢病毒构建稳定转染细胞系的步骤:[/b][/font][font=宋体] [/font][font=宋体][font=Calibri]1[/font][font=宋体]、选择慢病毒载体: 选择适当的慢病毒载体,通常是一个包含[/font][font=Calibri]LTR[/font][font=宋体]、包装信号、引导[/font][font=Calibri]RNA[/font][font=宋体]序列和多功能质粒载体的质粒。这个载体应该包含要表达的外源基因。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]2[/font][font=宋体]、转染慢病毒包装细胞: 使用慢病毒包装细胞系,例如[/font][font=Calibri]293T[/font][font=宋体]或其他适合的细胞系。这些细胞通常被选择因为它们能够支持慢病毒复制和包装。将慢病毒载体与包装蛋白的表达质粒一同转染进这些细胞中。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]3[/font][font=宋体]、病毒产生和收集: 慢病毒包装细胞会开始产生慢病毒颗粒,这些颗粒包含了慢病毒载体和外源基因。培养一定时间后,收集细胞培养上清液,这是富含病毒的液体。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]4[/font][font=宋体]、测定病毒滴度: 对采集的上清液进行病毒滴度的测定,通常可以通过转染一定数量的目标细胞,然后测定这些细胞的感染率来确定病毒的滴度。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]5[/font][font=宋体]、转染目标细胞: 将上一步获得的病毒用于转染目标细胞。这些目标细胞可以是要建立稳定转染细胞系的细胞。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]6[/font][font=宋体]、筛选稳定细胞系: 添加适当的筛选物质,例如抗生素,以选择表达了外源基因的细胞。这可以通过在培养基中添加抗生素,使得只有表达了外源基因的细胞能够存活下来。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]7[/font][font=宋体]、单克隆分离: 对稳定表达细胞群进行单克隆分离,以确保每个克隆都来自单一细胞。这有助于保持表达的一致性。[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]8[/font][font=宋体]、验证表达: 对所得的单克隆细胞系进行验证,确认外源基因的表达水平和稳定性。这可以通过[/font][font=Calibri][url=https://insevent.instrument.com.cn/t/jp][color=#3333ff]PCR[/color][/url][/font][font=宋体]、[/font][font=Calibri]Western blotting[/font][font=宋体]等分子生物学技术来实现。[/font][/font][font=宋体] [/font][font=宋体]通过这些步骤,可以建立一个稳定表达外源基因的慢病毒转染细胞系,为后续的实验和研究提供了有力的工具。这种方法常用于基因功能研究、药物筛选和基因治疗等领域。[/font][font=宋体] [/font][font=宋体][b]慢病毒构建稳转细胞系的优点:[/b][/font][font=宋体] [/font][font=宋体]与常用的转染方法相比,慢病毒构建稳转细胞系有以下几个优点:[/font][font=宋体] [/font][font=宋体]①高效性:慢病毒能够将外源基因整合到宿主细胞基因组中,实现稳定的外源基因表达。[/font][font=宋体] [/font][font=宋体]②特异性:由于慢病毒的感染和复制比较特异,只会影响一定类型的细胞,因此可以实现对具体细胞的选择性转染。[/font][font=宋体] [/font][font=宋体]③安全性:慢病毒的基因转移速度较缓慢,对宿主细胞和人体的损伤较小,因此具有较高的安全性。[/font][font=宋体] [/font][font=宋体][font=宋体]义翘神州提供[url=https://cn.sinobiological.com/services/stable-cell-line-development-service][b]稳转细胞株构建服务[/b][/url],包含过表达细胞系构建服务和[/font][font=Calibri]CHO[/font][font=宋体]稳定细胞株开发服务,详情可以关注:[/font][font=Calibri]https://cn.sinobiological.com/services/stable-cell-line-development-service[/font][/font]

◆产品特点 1、真正高效全自动:全球独家全自动多动能制片染色一体化设备,标本处理、制片、染色一次完成。 2、杜绝拖带污染:使用一次性加样针脱针系统和自动独立滴染湿式染色系统,杜绝了传统染色可能造成的交叉污染。 3、提取黏液包裹细胞:采用梯度离心分离及红细胞处理裂解和黏液消化技术三合一有效提取细胞及诊断成份,富集细胞及诊断成份,保证诊断细胞不丢失。 4、捕获病变细胞:根据人体不同类型细胞比重不同的特点,尤其是病变细胞比重大、沉降速度快,从而最大程度地捕获病变细胞和具诊断价值的成份,提高检出率。 5、薄层细胞均匀分布:基液使细胞均匀悬浮,保证随机性,任意取样涂片都具有代表性,形成均匀分布的真正薄层细胞涂片。 6、无需前处理:直接上机,标本无需前处理,三合一独家技术,细胞结构保存更完好,操作更加方便,省时高效,较国内外同类方法机型自动化程度更高。 7、高品质诊断保障:三合一独家技术有效提取细胞及诊断成份,完全清除黏液、红细胞等干扰成份,有利于病变细胞的鉴别诊断。 8、强大而简捷微机界面:人机对话式中文界面,可选择妇科及非妇科,不同数量及不同染色方法,操作更加方便,功能强大 9、绿色环保:不含一点甲醛,对于临床一线操作人员身体没有损害,无需采取特殊的防护。http://ng1.17img.cn/bbsfiles/images/2011/06/201106231242_301156_2324710_3.jpg
鸡蛋,在如今的社会里,更多时候是作为一种营养丰富的食品出现在我们的餐桌上。现代化大型养殖场如生产产品般输出鸡蛋的方式颠覆了人们对鸡蛋的认识,或许已经很少有人能够联想到从蛋黄蛋白到一个小生命的奇迹升华。但在人类漫长的历史中,农业是文明的核心。就在不太遥远的过去,大多数人还可以在家中目睹鸡生蛋、蛋生鸡的奇迹。这种神秘的现象让古时的人们感到好奇、困惑,甚至产生莫名的崇拜。我们华夏文明由雏鸡的诞生联想到世界的起源,“天地混沌如鸡子,盘古生其中”,看来在我们祖先的眼中,鸡蛋的孵化犹如天地诞生般神秘。这种“卵生崇拜”在史籍中屡见不鲜,如《史记·殷本纪》记述商朝人先祖契的来历时提到有娀氏的女儿简狄“见玄鸟坠其卵,简狄取吞之,因孕生契”,同样,在《史记·秦本纪》中,文章伊始就记载了颛顼的孙女女修织布时“玄鸟陨卵,女修吞之,生子大业”,而这位大业就是秦人的先祖。不得不佩服古人的想象,这玄鸟蛋孵化出了两个重要朝代。在漫长的历史中,这种对蛋朦胧而浪漫的崇拜逐渐融入了我们的文化中,直到如今,染红壳的鸡蛋依旧是新婚、生子、满月时,人们表达祝福的重要载体。随着科技的进步,人们对蛋的理解逐渐清晰,现在很多人都知道蛋和卵细胞有千丝万缕的关系。可是鸡蛋到底是否就是一个细胞?答案可谓五花八门,有人说整个鸡蛋就是一个放大的卵细胞,蛋壳内的那层膜是细胞膜,蛋清是细胞质,蛋黄是细胞核;也有人说蛋黄是卵细胞,卵黄膜就是细胞膜,蛋黄就是细胞质,而蛋黄上面的小白点是细胞核;还有人认为鸡蛋本就是由很多细胞构成的。
报告基因(reporter gene)是一种编码可被检测的蛋白质或酶的基因,是一个其表达产物非常容易被鉴定的基因。把它的编码序列和基因表达调节序列相融合形成嵌合基因,或与其 它目的基因相融合,在调控序列控制下进行表达,从而利用它的表达产物来标定目的基因的表达调控,筛选得到转化体。作为报告基因,在遗传选择和筛选检测方面必须具有以下几个条件:(1)已被和全序列已测定;(2)表达产物在受体细胞中不存在,即无背景,在被转染的细胞中无相似的内源性表达产物;(3)其表达产物能进行定量测定。在植物基因工程研究领域,已使用的报告基因主要有以下几种: 胭脂碱合成酶基因(nos)、章鱼碱合成酶基因(ocs)nos、ocs这两个基因是致瘤土壤农杆菌(Agrobacterium tumfaciens)的Ti质粒特有的,对Ti质粒进行改造,用相应的致瘤农杆菌转化植物体时,如果外源基因转入植物体中,则这两种报告基因在植物根茎 叶中均能表达,不受发育调控,检测时直接用转化体提取液进行纸电泳,染色后在紫外光下观察荧光即可。新霉素磷酸转移酶基因(nptⅡ)、氯霉素乙酰转移酶基因(cat) nptⅡ、cat及庆大霉素转移酶基因,均为抗生素筛选基因,相关的酶可以对底物进行修饰(磷酸化、乙酰化等),从而使这些抗生素失去对植物生长的抑制作 用,使得含有这些抗性基因的转化体能在含这些抗生素的筛选培养基上正常生长,也可以用转化体提取液体,外用同位素标记,放射自显影筛选转化体。氯霉素乙酰 转移酶基因测时可通过放射自显影观察。荧光素酶基因(luciferase Gene)1985年从北美荧火虫和叩头虫cDNA文库中出来的,该酶在有ATP、Mg2+、O2和荧光素存在下发出荧光,这样就可用植物整株或部分直接用X-光片 或专门仪器进行检测。具有检测速度快、灵敏度比cat基因高30~1000倍、费用低、不需使用放射性同位素等优点,得到了广泛的采用。β-D-葡萄糖苷酶基因该酶催化底物形成β-D-葡萄糖苷酸,它在植物体中几乎无背景,组织化学检测很稳定,可用分光光谱 、荧光等进行检测。除此之外还有庆大霉素转移酶基因等。在动物基因表达调控的研究中,已使用的报告基因主要有以下几种: 绿色荧光蛋白(gfp)基因等。绿色荧光蛋白来源于海洋生物水母,其基因可在异源组织中表达并产生荧光,GFP Cdnad 开放阅读框架长度约740bp,编码238个氨基酸残基,其肽链内部第65-67位丝氨酸-脱氢酪氨酸-甘氨酸通过自身环化和氧化形成一个发色基因,在长 紫外波长或蓝光照射下发出绿色荧光。转染后的细胞可在荧光显微镜或流式细胞仪(FACS)中直接观察基因的表达。此外还有β-半乳糖苷酶基因、二氢叶酸还原酶基因、氯霉素乙酰转移酶基因(cat)等。
请问各位高手,细胞培养和提取,特别是提取细胞液和细胞核都有哪些常用的方法啊?本人以前从未做过生物方面,麻烦说得详细、通俗一些,先行谢过了[em23] [em20] [em61]
美国叶史瓦大学艾伯特-爱因斯坦医学院研究人员发现细胞利用第一个已知的机制控制mRNA的存活。这些关于mRNA的发现可能对逆转癌症不受调控的细胞分裂提供启示。2011年12月22日,该研究发表在《细胞》期刊上。该研究通信作者Robert Singer博士说,“我们研究的mRNA分子命运类似希腊悲剧。它们的命运在诞生那一刻就被决定了。”该研究是利用Singer博士之前开发出的高级显微镜技术在酵母细胞中开展的,该技术也是第一次允许科学家在单个细胞中实时观察单个分子。制造蛋白的指令编码在基因的DNA序列上,而基因则是位于每个细胞核染色体中。但是要制造蛋白,基因的DNA编码必须拷贝或者说转录到mRNA分子上,然后mRNA从细胞核迁移到细胞蛋白制造工厂所在的细胞质。因为一旦mRNA存在,它就能够作为模板制造蛋白。因此科学家长期以来就怀疑当一种蛋白水平积累到危害的程度时细胞必须存在降解mRNA的方法。Singer博士说,“细胞在这个时候会以某种方式摧毁的mRNA,但是没有人知道这是如何发生的。”在他们寻求这种机制时,Singer博士和他的同事们集中注意在两种基因SWI5和CLB2,它们编码的蛋白调节细胞周期---细胞分裂期间复杂的一系列步骤,首先复制它的遗传物质,然后将遗传物质均匀地分配到两个子细胞中。为了合适地规划细胞周期,SWI5和CLB2基因编码的蛋白水平必须得到精致控制,这就意味着这两种基因制造的mRNA将是有目的降解的首要候选物。引人注目的是,研究人员发现这些mRNA事实上携带着最终将自己摧毁的分子“自我摧毁定时器(self-destruct timer)”。当基因被转录时,称作启动子区域的基因部分起着打开基因的作用,这样DNA将被拷贝到mRNA上。这些艾伯特-爱因斯坦医学院研究人员发现SWI5和CLB2启动子区域也有其他作用:它们招募一种蛋白 Dbf2p,这样当mRNA分子被合成时,Dbf2p就与它们结合。这些mRNA--由 SWI5和CLB2基因转录而来而且携带Dbf2p蛋白---就使得它们从细胞核运输到细胞质。在细胞质中,蛋白Dbf20p通过与Dbf2p连接在一起以便搭上mRNA分子,而且这两种蛋白一起就导致这些mRNA分子快速降解。Singer说,“我们的发现表明蛋白水平必须得到仔细控制,制造蛋白的基因含有启动子区域,正是该启动子区域在mRNA分子刚产生的时候就决定着它们死亡的命运,而且启动子区域是通过招募蛋白Dbf2p---mRNA合成和它的最终降解之间常见因子---来标记新制造的mRNA来行使的。 Dbf2p在mRNA诞生开始就与mRNA附着在一起,然后在接收到指示不应当制造更多蛋白的信号后作出应答,从而下达摧毁mRNA的命令。”Singer说,尽管这些观察都是与酵母细胞相关,但是他相信这种控制人mRNA降解的过程“将也是非常类似的”,可能用于对抗癌症。他注意到,“人们一旦获得对控制细胞周期和细胞分裂的机制新认识,就可以提出针对性的治疗方法来调节癌症中不受控制的细胞分裂。”
[font=宋体][font=宋体]在生物学和医学研究中,细胞增殖是一个关键过程,对于理解生命活动的基本规律以及疾病的发病机理具有重要意义。随着科技的发展,流式细胞仪作为一种高效、灵敏的分析工具,广泛应用于细胞增殖的检测。流式细胞仪通过快速分析单个细胞,可以对细胞周期、细胞增殖活性、细胞凋亡等多个方面进行研究。本文将探讨流式细胞仪在检测细胞增殖方面的主要方法,包括但不限于溴脱氧尿苷([/font][font=Calibri]BrdU[/font][font=宋体])掺入法、细胞周期蛋白检测法以及细胞大小分析法等,以期为读者提供全面的技术应用概览。流式细胞仪检测细胞增殖方法:[/font][/font][b][font=宋体][font=Calibri]1[/font][font=宋体]、[/font][font=Calibri]3H[/font][font=宋体](氚离子)掺入法[/font][/font][/b][font=宋体][font=宋体]原理:是在细胞[/font][font=Calibri]DNA[/font][font=宋体]合成时,用[/font][font=Calibri]3H[/font][font=宋体]脱氧胸腺嘧啶核苷代替普通的脱氧胸腺嘧啶核苷掺入新合成的[/font][font=Calibri]DNA[/font][font=宋体]中,增殖的细胞因为掺入[/font][font=Calibri]3H[/font][font=宋体]而具有放射性,通过定量检测样品细胞的放射性大小而反映细胞的增值活性[/font][/font][font=宋体][font=宋体]缺点:[/font][font=Calibri]1[/font][font=宋体])使用的是具有放射性的同位素,操作较为复杂,同时需要采取放射性保护措施 [/font][font=Calibri]2[/font][font=宋体])低比例高活跃增殖和高比例低活跃增殖可能得到的是相同的结果,用此方法无法进行鉴别 [/font][font=Calibri]3[/font][font=宋体])此方法无法进一步得到具有活性的增值细胞用于下一步的研究 [/font][font=Calibri]4[/font][font=宋体]) 此方法时间较短,无法检测加入前细胞的增殖情况,而且检测到放射性只能说明细胞[/font][font=Calibri]DNA[/font][font=宋体]合成,而不能提供合成[/font][font=Calibri]DNA[/font][font=宋体]的细胞是否进入增殖阶段的信息[/font][/font][b][font=宋体][font=Calibri]2[/font][font=宋体]、相对计数法[/font][/font][/b][font=宋体]原理:将对照组和各实验组控制在相同条件下直接计数然后比较计数结果得到增殖结论[/font][font=宋体]注意点:[/font][font=宋体][font=宋体]对照组与实验组每种细胞所加浓度必须相同,每组至少设置[/font][font=Calibri]3[/font][font=宋体]个复孔,这样每个孔可以得到[/font][font=Calibri]1[/font][font=宋体]个细胞数,将[/font][font=Calibri]3[/font][font=宋体]个复孔取平均值后就是这个组的结果。如果同时需要得到每孔目标细胞增殖后的绝对参数,在每孔细胞中加入[/font][font=Calibri]1*105PE[/font][font=宋体]标记的人工微球作为内参[/font][/font][font=宋体] [/font][font=宋体][font=宋体]收集各组的细胞于[/font][font=Calibri]EP[/font][font=宋体]管中,注意必须尽量将各组的所有细胞都收集起来。标记需要计数细胞的标志表型的荧光素偶联抗体,[/font][font=Calibri]4[/font][font=宋体]℃静置[/font][font=Calibri]30min[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]PBS[/font][font=宋体]洗涤一次,洗去游离的抗体[/font][/font][b][font=宋体][font=Calibri]3[/font][font=宋体]、示踪染料标记法[/font][/font][/b][font=宋体][font=宋体]示踪染料与细胞结合的方式:[/font][font=Calibri]1[/font][font=宋体])能够与细胞内的蛋白质上的氨基发生非特异性的共价结合 [/font][font=Calibri]2[/font][font=宋体])能够非特异性地嵌入细胞膜的脂质双分子层中与细胞发生非共价性结合[/font][/font][font=宋体] [/font][font=宋体][font=宋体]原理:示踪染料的荧光信号都很强,当细胞分裂时,母细胞内的染料会被平均分配到子细胞中,细胞荧光信号会被减弱一半,所以通过检测减弱的、发射示踪染料荧光信号的细胞比例就可以判断细胞增殖的强弱。当荧光强度减弱到标记时的[/font][font=Calibri]1/2[/font][font=宋体]以及以下的细胞都是增殖后的细胞,这些细胞所占比例越高则代表细胞增殖越活跃[/font][/font][font=宋体] [/font][font=宋体]标记方法:[/font][font=宋体][font=宋体]①纯化增殖反应的目标细胞,将细胞的浓度调整为[/font][font=Calibri]1*106/ml[/font][font=宋体],加入[/font][font=Calibri]CFSE[/font][font=宋体],其标记浓度为[/font][font=Calibri]5[/font][font=宋体]微摩尔[/font][font=Calibri]/[/font][font=宋体]升。置于[/font][font=Calibri]37[/font][font=宋体]℃水浴中标记[/font][font=Calibri]15min[/font][font=宋体],在标记过程中每隔一段时间混匀细胞一次[/font][/font][font=宋体] [/font][font=宋体][font=宋体]②加入预冷、含有血清的培养基终止标记,在[/font][font=Calibri]4[/font][font=宋体]℃冰箱中静置[/font][font=Calibri]5min[/font][font=宋体],离心沉淀[/font][/font][font=宋体] [/font][font=宋体][font=宋体]③用培养基再洗涤一次,尽量洗净未结合的游离的[/font][font=Calibri]CFSE[/font][font=宋体],然后将目标细胞静置在增殖体系中[/font][/font][font=宋体] [/font][b][font=宋体][font=Calibri]4[/font][font=宋体]、[/font][font=Calibri]BrdU[/font][font=宋体]和[/font][font=Calibri]EdU[/font][font=宋体]掺入法[/font][/font][/b][font=宋体][font=Calibri]BrdU[/font][font=宋体]:[/font][font=Calibri]5-[/font][font=宋体]溴脱氧尿嘧啶核苷是胸腺嘧啶核苷的类似物,其特点是胸腺嘧啶环上[/font][font=Calibri]5[/font][font=宋体]位[/font][font=Calibri]C[/font][font=宋体]连接的甲基被溴取代,在细胞增殖[/font][font=Calibri]DNA[/font][font=宋体]合成时可以与内源性的胸腺嘧啶核苷竞争掺入到新合成的[/font][font=Calibri]DNA[/font][font=宋体]中,而[/font][font=Calibri]BrdU[/font][font=宋体]抗体可以特异性的识别[/font][font=Calibri]BrdU[/font][font=宋体],不与胸腺嘧啶核苷结合,所以可以用于检测细胞增殖[/font][/font][font=宋体][font=宋体]适用范围:适用于体内检测目标细胞的增殖,一般将[/font][font=Calibri]BrdU[/font][font=宋体]掺入小鼠的应用水中或经腹腔注射,经过一段时间后,取出目标细胞制成单细胞悬液然后用多聚甲醛固定细胞,后用打孔剂皂苷在细胞膜上打孔,最后标记荧光素偶联抗[/font][font=Calibri]BrdU[/font][font=宋体]抗体,目标细胞的[/font][font=Calibri]BrdU[/font][font=宋体]阳性细胞就是增殖的细胞,阳性比例越高,增殖越活跃。[/font][/font][font=宋体] [/font][b][font=宋体][font=Calibri]5[/font][font=宋体]、其他方法[/font][/font][/b][font=宋体][font=宋体]细胞周期法检测细胞增殖:流式细胞术能够检测细胞内[/font][font=Calibri]DNA[/font][font=宋体]的含量,所以可以检测细胞周期。处于[/font][font=Calibri]S[/font][font=宋体]期的细胞,[/font][font=Calibri]DNA[/font][font=宋体]的量处于二倍体和四倍体之间[/font][font=Calibri] [/font][font=宋体]处于[/font][font=Calibri]G2/M[/font][font=宋体]期时,[/font][font=Calibri]DNA[/font][font=宋体]量为四倍体。处于[/font][font=Calibri]S[/font][font=宋体]期和[/font][font=Calibri]G2/M[/font][font=宋体]期的细胞比例越高说明细胞增殖越活跃[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]PCNA[/font][font=宋体]检测细胞增殖:[/font][font=Calibri]PCNA[/font][font=宋体](增殖细胞核抗原),在细胞核合成且只存在于细胞核内,是[/font][font=Calibri]DNA[/font][font=宋体]聚合酶的辅助蛋白,所以与细胞[/font][font=Calibri]DNA[/font][font=宋体]的合成关系密切,是反映细胞增殖状态的良好指标[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]Ki-67[/font][font=宋体]检测细胞增殖:是一种与细胞增殖特异相关的核抗原[/font][/font][font=宋体] [/font][font=宋体][font=Calibri]CD71[/font][font=宋体]检测细胞增殖:是转铁蛋白受体,表达于细胞的表面,该受体广泛表达于各种恶性肿瘤细胞表面,正常细胞表达较少,与肿瘤细胞的增殖密切相关[/font][/font][font=宋体] [/font][font=宋体][font=宋体]义翘神州提供[url=https://cn.sinobiological.com/services/flow-cytometry-service][b]流式细胞检测技术服务[/b][/url],更多关于流式细胞仪检测细胞增殖详情欢迎咨询,详情可以关注:[/font][font=Calibri]https://cn.sinobiological.com/services/flow-cytometry-service[/font][/font][b][font=宋体] [/font][font=宋体][font=宋体]义翘神州:蛋白与抗体的专业引领者,欢迎通过百度搜索[/font][font=宋体]“义翘神州”与我们取得联系。[/font][/font][/b]
一、实验目的1、掌屋凋亡细胞的形态特征 2、学会用荧光探针对细胞进行双标记来检测正常活细胞、凋亡细胞和坏死细胞的方法二、实验原理 细胞死亡根据其性质、起源及生物学意义区分为凋亡和坏死两种不同类型。凋亡普遍存在于生命界,在生物个体和生存中起着非常重要的作用。它是细胞在一定生理条件下一系列顺序发生事件的组合,是细胞遵循一定规律自己结束生命的自主控制过程。细胞凋亡具有可鉴别的形态学和生物化学特征。在形态上可见凋亡细胞与周围细胞脱离接触,细胞变园,细胞膜向内皱缩、胞浆浓缩、内质网扩张、细胞核固缩破裂呈团块状或新月状分布、内质网和细胞膜进一步融合将细胞分成多个完整包裹的凋亡小体,凋亡小体最后被吞噬细胞吞噬消化。在凋亡过程中细胞内容物并不释放到细胞外,不会影响其它细胞,因而不引起炎症反应。在生物化学上,多数细胞凋亡的过程中,内源性核酸内切酶活化,活性增加。核DNA随机地在核小体的连接部位被酶切断,降解为180-200bp或它的整倍数的各种片断。如果对核DNA进行琼脂糖电泳,可显示以180-200bp为基数的DNA ladder(梯状带纹)的特征。相比之下,坏死是细胞处于剧烈损伤条件下发生的细胞死亡。细胞在坏死早期即丧失质膜完整性,各种细胞器膨胀,进而质膜崩解释放出其中的内容物,引起炎症反应,坏死过程中细胞核DNA虽也降解,但由于存在各种长度不等的DNA片断,不能形成梯状带纹,而呈弥散状。一些温和的损伤刺激及一些抗肿瘤药物可诱导细胞凋亡,通常这些因素在诱导凋亡的同时,也可产生细胞坏死,这取决于损伤的剧烈程度和细胞本身对刺激的敏感程度。三尖杉酯碱(HT)是我国自行研制的一种对急性粒细胞白血病,急性单核白血病等有良好疗效的抗肿瘤药物。研究表明HT在0.02~5μg/ml范围内作用2小时,即可诱导HL-60细胞凋亡,并表现出典型的凋亡特征。本实验用1μg/ml HT在体外诱导培养的HL-60细胞发生凋亡,同时也有少数细胞发生坏死。用Hoechst33342和碘化丙啶(propidium iodide,PI)对细胞进行双重染色,可以区别凋亡、坏死及正常细胞。细胞膜是一选择性的生物膜,一般的生物染料如PI等不能穿过质膜。当细胞坏死时,质膜不完整,PI就进入细胞内部,它可嵌入到DNA或RNA中,使坏死细胞着色,凋亡细胞和活细胞不着色。而一些活细胞染料由于为亲脂性物质,可跨膜进入活细胞,因而可进行活细胞染色。Hoechst33342是一种活性荧光染料且毒性较弱,它是双苯并咪唑的一种衍生物。与DNA特异结合(主要结合于A-T)碱基区),显示凋亡细胞和活细胞。凡是看到有凋亡小体的细胞都是凋亡细胞。三、实验用品1、试剂:三尖杉酯碱(HT),300μg/ml,100mmol/L Tris-HCl(pH7.5),5mol/L EDTA缓冲液、碱性裂解液:0.2mol/L NaOH, 1%SDS、醋酸钠:3mol/L KAc(pH4.8);异丙醇;70%乙醇;溴酚蓝,蔗糖指示剂。TBE电泳缓冲液,1%琼脂糖,溴乙锭。PI母液:500μg/ml;Ho33342母液:2mmol/L。 2、仪器设备:荧光显微镜,电泳仪,电泳槽,微量加样器(1ml,100μl)0.5、1.5ml离心管,载玻片,盖玻片。四、实验材料人早幼粒白血病HL-60细胞,用含10%小牛血清的RPMI1640培养基在37℃,5%CO2条件下培养。五、方法步骤 1、三尖杉酯碱诱发HL-60细胞凋亡 (1)实验前约24小时,接种两瓶HL-60细胞,标记①、②,每瓶含约6ml培养液,置37℃,5%CO2培养箱培养。 (2)实验前约2.5小时,当细胞密度达到70%,①号瓶加入三尖杉酯碱200μl,使终浓度为1μg/ml,②号瓶中加入同等量PBS(pH7.4)作对照。共同放入培养箱中继续培养2.5小时。2、Ho33342和PI双重染色鉴别三种细胞 (1)染色:将瓶中的细胞摇匀取200μl于1.5ml的离心管中,加入Ho33342母液2μl,PI 20μl,染色15分钟。 (2)滴片:取一载玻片用双面胶围成一小室,从离心管中各取以上染色后的细胞悬液10μl,加入小室内盖上盖玻片,荧光镜下用紫外激发光,高倍镜下观察,区别三种细胞,并注意三者比例。六、注意事项1、诱导培养HL-60细胞时间要准确; 2、荧光显微镜下观察细胞时,由于荧光易碎灭,观察时要尽量快。
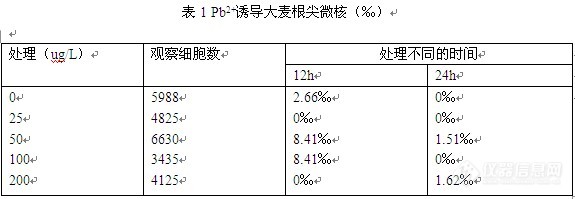
[color=#d40a00][size=3][size=2]维权声明:本文为xiaodaren原创作品,本作者与仪器信息网是该作品合法使用者,该作品暂不对外授权转载。其他任何网站、组织、单位或个人等将该作品在本站以外的任何媒体任何形式出现的,均属侵权违法行为,我们将追究法律责任。[/size]原创大赛又来啦!!今年一不留神错过了7月的礼品!还好8月奖品更加精彩,赶紧小跑着来发原创!!!呵呵这次与大家分享的其实是一个很简单的实验过程,但是简单不乏味,从这次试验过程中我也学到了一些东西,所以拿出来与各位一同分享!!!看到这次奖品还可以许愿~~~真是人性化啊!!!正好我们开始租房子自己做饭啦~~所以想要一个高压锅炖排骨~~~不知道可不可以,价值嘛就无所谓啦!好啦,闲言碎语就不说啦~~~下面实验过程华丽丽登场~~~~[/size][/color][color=#0162f4] 重金属对植物的毒害作用日前己知是多方而的。在生理生化方而的毒去表现在使烟草叶绿素a. b含量下降 羊角月芽藻叶绿素a含量下降,膜通透性加大、过氧化物酶活性增加 小白菜抗坏血酸含量下降等。这些都充分表明铅使植物光合作用降低、加速了过氧化衰老速度。然而重金属铅以其化合物(pbc12)的形式普遍存在于土壤、大气和水中。近些年来,我国的采业、冶金业以及交通运输业的发展突飞猛进.铅污染日益严重,对人类、农业、环境造成极大的危重。铅不是生物生存所必需的元素而属于有害元素,并且进入生物体内很难排出使富积下来毒害机体。因此.严防铅中毒己成为环境质量控制的一个内容铅对人类和动物的毒害作用主要是对神经、免疫系统及造血_功能的毒害。对植物的毒害主要表现在细胞遗传学。生理生化等代谢方而同时根尖微核及染色体畸变技术己广泛应用于植物毒理学研究发挥了重要作用。同时该项技术己作为一项世界性的生物学检测指标。[/color][color=#0162f4] 遗传损伤主要研究以下几个指标:(一)微核,简称mcn,是真核类生物细胞中心的一种异常结构,一般认为它是由有丝分裂后期丧失着丝粒的染色体片段产生。这些断片或染色体在分裂过程中行动滞后,分裂末期不能进入主核,形成主核之外的核块。当细胞进入下一次分裂间期,他们便浓缩成主核之外的小核,即微核。微核大小在主核1/3以下,与主核分离,着色与主核一致或较深,呈圆形或椭圆形。(二)有丝分裂指数(MI)指观察细胞中处于有丝分裂相的细胞数。(三)根尖细胞染色体异常:断裂,滞后,粘连,核固缩。本文通过铅对大麦细胞遗传学毒害作用方而的一些研究.为农业旱期预测诊断铅对作物的危害去除提供了重要的理论依据。[/color] [b]1材料和方法[/b] 1.1 材料 1.1.1重金属处理液,1N HCl,石炭酸品红溶液 1.1.2.培养皿,显微镜,水浴锅,计数器,镊子,试管,载玻片,盖玻片 1.1.3大麦Hordeum Vulgare(2n=14) 1.2方法 1.2.1种子萌发、催芽 1.2.2重金属处理:根长至2~3cm后,开始处理,12h更换一次处理液,处理时间为12h、24h。染毒后蒸馏水恢复培养24h。 1.2.3固定 卡诺固定液固定24h。 1.2.4酸解 从固定液中取出根尖,蒸馏水洗静,用1N HCl 60℃酸解。 1.2.5染色 酸解后用蒸馏水洗涤两遍,根尖切下,加2~3滴染液进行染色。 1.2.6观察 每一处理观察6~8个根尖约3000个细胞。 [b]2.结果[/b] 2.1 Pb2+诱导对大麦根尖微核的影响(见表1):[img]http://ng1.17img.cn/bbsfiles/images/2017/01/201701191653_630965_1856701_3.jpg[/img] 由表1可见,不同染毒时间下,随着染毒浓度的增加(0.025一0.2 mg/L) 诱发大麦根尖微核率有所增加,但是随处理时间的增加(12h—24h)微核率有波动且其增长趋势不显著。 2.2 Pb2+诱导对大麦根尖有丝分裂指数的影响:[img]http://ng1.17img.cn/bbsfiles/images/2017/10/201008191045345109_01_1856701_3.jpg[/img] 表2资料看到:铅可以显著降低大麦根尖细胞的有丝分裂指数,对植物生长具有抑制作用,呈浓度依赖和时间依赖关系,随着铅浓度的增高,在同一处理时间下(如24h下不同浓度所造成的影响)其有丝分裂指数明显下降而在同一浓度下随着处理时间的增加根尖分生细胞有丝分裂指数都有减少的趋势。 2.3 Pb2+诱导对大麦根尖有丝分裂染色体的影响[img]http://ng1.17img.cn/bbsfiles/images/2017/10/201008191047069118_01_1856701_3.jpg[/img] 从表3实验资料看到.大麦根尖分生细胞在细胞分裂过程中,染色体的形态变化对铅的毒害更为敏感。出现了染色体桥、断裂和粘连及核固缩。随着浓度的增加处理时间的延长,波观察的4种畸变的绝对值都在增加。从四种畸变综合统计结果来看.铅浓度愈大、处理时间愈长畸变率愈高。凡是高浓度、长时间 (24 h)处理的大麦根尖分生细胞都停止了有丝分裂,不久都将死亡。 [b]3.讨论[/b] 3.1 Pb2+诱导对大麦根尖微核的影响 由表1可见,不同染毒时间下,随着染毒浓度的增加(0.025一0.2 mg/L) 诱发大麦根尖微核率有所增加,对植物具有遗传损伤效应,且高浓度短时间作用和低浓度长时间作用具有等效性,与阴性对照组有显著差异,随处理时间的增加(12h—24h)微核率有波动且其增长趋势不显著,此可能为实验不当所造成,其原因需经进一步探究。 3.2 Pb2+诱导大麦根尖分生细胞有丝分裂指数的影响 试验统计了Pb2+诱导在不同浓度处理及不同时间下对大麦根尖细胞有丝分裂的影响.以指示重金属对植物细胞的毒害作用。细胞分裂指数=(分裂细胞数/观察细胞总数)*100%. 表2资料看到:铅可以显著降低大麦根尖细胞的有丝分裂指数,对植物生长具有抑制作用,呈浓度依赖和时间依赖关系,随着铅浓度的增高,在同一处理时间下(如24h下不同浓度所造成的影响)其有丝分裂指数明显下降,说明高浓度的Pb2+可抑制有丝分裂的发生,另外,同一浓度下随着处理时间的增加根尖分生细胞有丝分裂指数都有减少的趋势。 3.3 Pb2+诱导对大麦根尖有丝分裂染色体的影响 从表3实验资料看到.大麦根尖分生细胞在细胞分裂过程中,染色体的形态变化对铅的毒害更为敏感。出现了染色体桥、断裂和粘连及核固缩。随着浓度的增加处理时间的延长,波观察的4种畸变的绝对值都在增加。从四种畸变综合统计结果来看.铅浓度愈大、处理时间愈长畸变率愈高。凡是高浓度、长时间 (24 h)处理的大麦根尖分生细胞都停止了有丝分裂,不久都将死亡。 正常根尖细胞具有稀疏的细胞质,并有均匀的染色质。经Pb2+诱导后有些根尖细胞核核浆减少,核体积缩小,核边增厚,核染色质局部或全部凝集,由网状至完全固缩而致密,着色很深,形成固缩核,与此同时细胞质变得稀少,部分细胞体积随之变小。Pb2+诱导后大麦根尖分生区细胞的核固缩统计结果见表3, 表3数据显示,Pb2+诱导细胞核固缩具有时间效应和剂量效应关系,随着Pb2+诱导浓度增大和作用时间延长,大蒜幼根细胞核固缩率增高。不同根尖细胞中固缩核数相差较大,有的根尖核固缩的细胞很多,有的根尖却很少固缩细胞。这种情形可能是由于几不同个体对环境的适应性不同,有的个体对Pb2+的作用适应较强,继续分裂生长 而有的个体不能适应Pb2+毒理作用,致使细胞停止分裂并逐步死亡。 3.4 综上所述,铅对大麦根尖分生细胞的影响主要表现为:第一,铅进入植物体后.主要集中在根部细胞核内.在组织间迁移率极低所以阻碍细胞向分化状态发展。铅对大麦根尖细胞有丝分裂指数表现出浓度与时间的叠加抑制效应。第二,铅能诱导染色体畸变。畸变率表现为浓度与时间的叠加诱导效应。第三,铅对大麦根尖分生细胞的细胞遗传学毒理从形态上看是染色体各种畸变的形式,实质上是对间期DNA复制前后的阻断、干扰和影响。 铅对细胞遗传学的毒去作用.主要表现为染色体畸变和微核效应。不少学者认为:这是因为抑制和干扰了G1期触发蛋白( trigger Protein)的合成,限制了由G1期进入S期。一但进入了S期,铅不能影响DNA复制和合成。因此,染色体畸变和微核的形成是S期DNA受到损伤的结果。关于铅对植物毒去的机理研究.日前仍处于资料积累阶段有待进一步深入探讨。[color=#d40a00]本来如果显微镜可以照相的话可以让大家看到我所观察的结果~~但是无奈设备无法达到要求啊~~~大家就只能看我的结果说明了~~~如果有所过同类实验的朋友们可以分享补充哈~~~~[/color]







 我要推广仪器
我要推广仪器
 下载APP
下载APP