推荐厂家
暂无
暂无
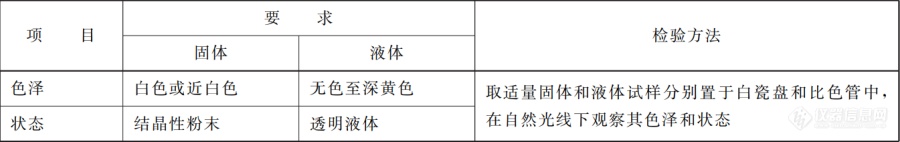
[font='calibri'] [/font][font='calibri'][size=14px] [/size][/font][size=29px]食品添加剂异构化乳糖[/size][font='calibri'][size=29px] [/size][/font][font='calibri'][size=21px] [/size][/font][font='等线'][size=21px]董堃河[/size][/font][font='黑体'][size=14px]摘要[/size][/font][font='黑体']:[/font][font='黑体']异构化乳糖是以乳糖为原料通过异构化作用制得的一种半合成双糖,具有益生元的作[/font][font='黑体']用特点。近年来,食品工业尤其是乳制品工业中益生元的应用已经成为了趋势。异构化乳糖[/font][font='黑体']是以乳糖为原料,由乳糖的葡萄糖部分在碱性条件下异构为果糖而形成的双糖。异构化乳糖[/font][font='黑体']因其对人体的有益作用而广泛应用于医药、营养和食品工业。异构化乳糖是一种双糖益生元,[/font][font='黑体']有效增殖肠道内的双歧杆菌菌群。它在食品领域中应用时添加量比较低且容易检测;又因其[/font][font='黑体']能减少肠道运转时间和降低体内氨含量,异构化乳糖被全球用于治疗慢性便秘和肝性脑病等[/font][font='黑体']医疗行业。鉴于国外对异构化乳糖的研究与应用一直未停步并为了提高我国消费者和 研发[/font][font='黑体']人员对于异构化乳糖的认知程度,本文从异构化乳糖的[/font][font='黑体']理化性质,应用,检测,限量,标准[/font][font='黑体']出发,综述了其常见的生理功能及在食品工业中的应用现状。[/font][font='黑体'][size=16px]关键词[/size][/font][font='黑体']:异构化乳糖,理化性质,应用,检测,限量,标准[/font][font='黑体']异构化乳[/font][font='黑体']糖即[/font][font='黑体']4-O-β-吡喃半乳糖-D-果糖[/font][font='黑体'],[/font][font='黑体']也被称为乳果糖或乳酮糖,是由半乳糖和果糖经 β-(1 → 4)糖苷键连接的双糖。它不能自然产生,是使用不同的催化剂(化学试剂、酶制剂或电激活)由乳糖发生异构化而产生的[/font][font='黑体']。[/font][font='黑体']研究表明,异构化乳糖具有促进双歧杆菌增殖[/font][font='黑体'][[/font][font='黑体']1],改善肠道菌群,防止便秘,增强机体免疫,促进人体对钙、镁的吸收等作用[/font][font='黑体'][[/font][font='黑体']2],因此异构化乳糖可广泛用于保健食品中,可添加到乳饮料、碳酸饮料、果汁饮料、糖果、乳粉等食 品中作膳食疗效食品,也可单独作医疗用品[/font][font='黑体'][[/font][font='黑体']3][/font][font='黑体']。[/font][font='黑体']我国GB 2760-2014《食品安全国家标准食品添加剂使用标准》中规定异构化乳糖液可应用于乳粉(包括加糖乳粉)和奶油[/font][font='黑体']粉及其[/font][font='黑体']调制产品、饼干、婴幼儿配方食品和饮料(包装饮用水除外)。[/font][font='黑体']目前[/font][font='黑体'],异构化乳糖在中老年奶粉中已经有所应用,但是在其他食品行业中的开发应用还很少。[/font][font='黑体'][[/font][font='黑体']4]本文着重介绍异构化乳糖的生理功能及[/font][font='黑体']应用、检测及限量标准,[/font][font='黑体']以期为科研工作者开发异构化乳糖产品提供参考[/font][font='黑体']。[/font][font='等线'][size=13px]1. [/size][/font][font='等线'][size=13px]异构化乳糖液的理化性质[/size][/font][font='等线'][size=13px]异构化乳糖是乳糖的异构化产物,因此它们具有相同的分子式[/size][/font][font='等线'][size=13px]C[/size][/font][font='等线'][size=13px]12[/size][/font][font='等线'][size=13px]H[/size][/font][font='等线'][size=13px]22[/size][/font][font='等线'][size=13px]O[/size][/font][font='等线'][size=13px]11[/size][/font][font='等线'][size=13px]和分子量[/size][/font][font='黑体'](Mr=342.3)。碱金属类氢氧化物和硼酸通常用来催化该异构化反应。异构化乳糖是由乳糖[/font][font='黑体']的葡萄糖部分异构为果糖而形成的双糖,且半乳糖和果糖以β-(1 → 4[/font][font='黑体'])[/font][font='黑体']糖苷键连接。商[/font][font='黑体']业生产异构化乳糖一般以氢氧化钠为催化剂,在一定条件下使乳糖的末端葡萄糖发生异构化[/font][font='黑体']反应转化成果糖而[/font][font='黑体']得(见图1)。[/font][img]https://ng1.17img.cn/bbsfiles/images/2021/07/202107262012362007_2338_1608728_3.jpeg[/img][font='黑体']图1[/font][font='黑体'] [/font][font='黑体'] 异构化乳糖 , 通常是淡黄色的透明的糖酱状糖液,有令人舒适的甜味[/font][font='黑体']。[/font][font='黑体']异构化乳糖的[/font][font='黑体']实测值是砂糖的60[/font][font='黑体']~[/font][font='黑体']70[/font][font='黑体']%。[/font][font='黑体']异构化乳糖的颜色是透明的淡黄色[/font][font='黑体'],[/font][font='黑体']长期保存或在高温下连续加[/font][font='黑体']热[/font][font='黑体'],[/font][font='黑体']异构化乳糖和果糖的着色程度相仿[/font][font='黑体']。[/font][font='黑体']因此在气温高的季节[/font][font='黑体'],[/font][font='黑体']最好储放在冷暗的地方[/font][font='黑体']。[/font][font='黑体']添[/font][font='黑体']加山梨醇等糖醇[/font][font='黑体'],[/font][font='黑体']可防止异构化乳糖色泽的加深[/font][font='黑体'][[/font][font='黑体']5][/font][font='黑体']。[/font][font='黑体']异构化乳糖粉末是白色到近白色无味结晶粉末(相对甜度0.6~0.8)[/font][font='黑体'],[/font][font='黑体']可溶于水,微[/font][font='黑体']溶于甲醇,不溶于乙醚,熔点介于168.5 和170 ℃。在30 ℃时它在水中的溶解度为76.4%[/font][font='黑体'](w/w),在90[/font][font='黑体']℃[/font][font='黑体']增加到86%。它的甜度为蔗糖的 0.48~0.62,比乳糖的甜度高1.5 倍。异[/font][font='黑体']构化乳糖经酸水解得到半乳糖和果糖。在低pH值条件下加热到 130 ℃10 min,异构化乳糖[/font][font='黑体']能保持稳定,很少被降解。这种相对较高的稳定性使得其能够满足正常食品加工的需要[6][/font][font='黑体']。[/font][font='黑体']具体理化性质见图2。[/font][align=center][img]https://ng1.17img.cn/bbsfiles/images/2021/07/202107262012366088_2515_1608728_3.jpeg[/img][/align][align=center][font='黑体']表1[/font][font='黑体'] 异构化乳糖液和粉末的主要成分[/font][font='黑体'][[/font][font='黑体']4][/font][/align][font='黑体'] [/font][font='黑体'] 以上是药典上的要求;异构化乳糖之外的其它糖的浓度通常低的多。成分上的差异在药学上 和药理学上不重要;在非常低的残糖浓度和/或高异构化乳糖浓度下,糖浆微生物会变得不稳定。[/font][font='黑体'] [/font][font='黑体'] 市场上的异构化乳糖有两种类型:异构化乳糖液和异构化乳糖结晶粉。异构化乳糖液[/font][font='黑体']是无色 或微黄色、无味和微甜的糖浆。其微黄和微甜可 能是由于糖浆中含半乳糖、乳糖、[/font][font='黑体']依匹乳糖、塔格糖和果糖引起的。异构化乳糖糖浆应储存在室温下。颜色改变或不透明可[/font][font='黑体']能是受热或见光引起的,但并不影响异构化乳糖的活性。粉状的异构化乳糖是白色、无味[/font][font='黑体']的结晶粉。异构化乳糖的理化性质见[/font][font='黑体']图2[/font][font='黑体']。[/font][align=center][img]https://ng1.17img.cn/bbsfiles/images/2021/07/202107262012367283_5497_1608728_3.jpeg[/img][/align][align=center][font='黑体']图2[/font][/align][font='黑体']与其底物乳糖相比,异构化乳糖中的 β-1,4糖苷键不能被人肠道消化酶所分解。虽[/font][font='黑体']然绝大多数白种人可以消化吸收乳糖,但不能吸收异构化乳糖,主要因为其是通过可分解糖[/font][font='黑体']的结肠细菌所代谢。异构化乳糖有益于乳酸菌的生长和代谢,而不利于许多水解蛋白的致病[/font][font='黑体']菌的生长,这种特征受生物、医学等多方面作用,表现出与其它益生元接近的效果。例如,[/font][font='黑体']乳糖醇同异构化乳糖具有相同的分子式和分子量,都不在小肠中分解,但二者对细菌代谢、[/font][font='黑体']粪便pH值和转运时间等参数的影响差异显著。已有研究表明乳糖醇在相同剂量下效果明显[/font][font='黑体']较差,且只有其环状部分才起到益生元作用,而醇基被认为是通过渗透方式发挥作用。[/font][font='黑体'] [/font][font='等线'][size=13px]2. [/size][/font][font='等线'][size=13px]异构化乳糖的应用[/size][/font][font='等线'][size=13px]异构化乳糖应用于食品行业已经50多年。 1991年日本政府为特殊保健用食品(FOSHU)[/size][/font][font='黑体']立法,异构化乳糖作为功能性配料允许添加到保健食品中。2010年,欧盟食品安全局 [/font][font='黑体'](EFSA)对异构化乳糖得出的结论是:在单份食物消耗下[/font][font='黑体'],[/font][font='黑体']消耗至少10g的异构化乳糖,能够建立“消耗异构化乳糖”和“减少肠道转运时间” 的因果关系[/font][font='黑体'][[/font][font='黑体']7]。异构化乳糖液是双歧杆菌的增殖因子,具有帮助消化吸收蛋白质、乳糖和产生维生素 B 组等功能;在我国批准使用并可以添加到乳粉、婴儿配方粉、饼干和饮料中等[/font][font='黑体']。[/font][font='黑体']异构化乳糖作为食品原料具有稳定性高,耐高温,耐酸,添加量低,良好的水溶性,较[/font][font='黑体']低的热量值,增强风味、不引起龋齿以及加工性能好[11]。 异构化乳糖是乳糖的差向异构[/font][font='黑体']体,甜度比乳糖高且溶解性好,可以用于甜味剂和矫味剂[[/font][font='黑体']1[/font][font='黑体']2]。1960年日本森永乳业在全[/font][font='黑体']球率先开发出添加异构化乳糖的婴幼儿配方奶粉。1995 年,R. Nagendra[13]等研究了含[/font][font='黑体']异构化乳糖的婴儿配方奶的稳定性。测试温度是 5℃,27℃和 38℃ ;测试时间 20w[/font][font='黑体'];[/font][font='黑体']产[/font][font='黑体']品马口铁罐包装和氮气密封。结果表明在任何温度下含异构化乳糖的奶粉和无异构化乳糖的[/font][font='黑体']奶粉在湿度和溶解度上没有变化。添加异构化乳糖并没有影响产品的可接受度而且在 38℃[/font][font='黑体']下能达到 16w 的保质期。[/font][font='黑体']临床上,异构化乳糖适用于10[/font][font='黑体']~[/font][font='黑体']30g 剂量的便秘的对症治疗和60[/font][font='黑体']~[/font][font='黑体']100g 剂量的门静脉系统脑病的治疗[/font][font='黑体'],[/font][font='黑体']但是当其作为一种功能性食品的成分时,使用的剂量往往低于10g[8][/font][font='黑体']。[/font][font='黑体']Melanie 等[8]采用了2[/font][font='黑体']~[/font][font='黑体']5g /d并持续5d的剂量的异构化乳糖[/font][font='黑体'],[/font][font='黑体']作用于TIM-2 系统,这[/font][font='黑体']是一种近端大肠的计算机控制模型,代表人来源的复杂、高密度、代谢活跃的 厌氧微生物[/font][font='黑体']群,结果发现,虽然 2g 的异构化乳糖已经增加了肠内容物的短链脂肪酸水平,但仍需要 [/font][font='黑体']5g 的异构化乳糖,来充分发挥出其益生元效应,即双歧杆菌、乳酸杆菌和厌氧菌的较高细[/font][font='黑体']菌计数,乙酸盐、丁酸盐和乳酸盐的增加,以及支链脂肪酸的减少、pH 值的下降和氨气的[/font][font='黑体']减少。另一方面,Tsunesuke T 等[9]研究发现,每天摄入 0. 65g 的异构化乳糖,能够[/font][font='黑体']起到促进成年人肠道内双歧杆菌的增殖、软化大便和改善肠道菌群的作用。Mizota 等[10][/font][font='黑体']通过临床试验研究了低剂量的异构化乳糖对成人肠道功能的影响,发现正常成人摄入异构化[/font][font='黑体']乳糖( 3g /d 和 5g /d) 后排便次数和粪便软化程度增加 还发现服用期 间粪便中双歧菌[/font][font='黑体']的比率由之前的 22. 4% 增加至 50. 5%,且粪便中腐败物的含量显著减少。[/font][img]https://ng1.17img.cn/bbsfiles/images/2021/07/202107262012368236_4210_1608728_3.jpeg[/img][font='黑体'] [/font][font='黑体']表2[/font][img]https://ng1.17img.cn/bbsfiles/images/2021/07/202107262012371212_9056_1608728_3.jpeg[/img][font='黑体']表3[/font][font='等线'][size=13px]3. [/size][/font][font='等线'][size=13px]异构化乳糖的检测[/size][/font][font='黑体'] [/font][font='黑体'] 目前,测定异构化乳糖的方法包括分光光度法[3]、液相-质谱法[14]、[url=https://insevent.instrument.com.cn/t/3p][color=#3333ff]离子色谱[/color][/url]法[15]、[/font][font='黑体']薄层色谱法[16]、液相色谱法[17]等。分光光度法前处理复杂结果易受干扰,检测需要多种[/font][font='黑体']酶试剂费用高,对人员要求较高;液相-质 谱法和[url=https://insevent.instrument.com.cn/t/3p][color=#3333ff]离子色谱[/color][/url]法定量准确,但是仪器价格昂贵[/font][font='黑体']或普及 率低,不便于日常监测;薄层色谱法灵敏度低,只能作半定量分析,不能准确定量[/font][font='黑体'];以上方法均不便在实际检测中应用。液相色谱检测异构化乳糖的方法具有准确、快速、成[/font][font='黑体']本低等特点,是目前检测异构化乳糖的理想的方法。[/font][font='黑体']同时[/font][font='黑体']酶法以其仪器成本低、操作简单,检测结果准确、稳定等优点,成为目前较为成熟且应用广泛的一种方法[3][/font][font='黑体']。[/font][font='黑体']对于液相色谱检测的色谱条件选择[[/font][font='黑体']17][/font][font='黑体']:[/font][font='黑体']选择粒径5 μm的氨基柱,调节流动相乙腈和水的比例,但出峰时间早,异构化乳糖[/font][font='黑体']和乳糖分不开,出峰时间晚可以分开但是峰型展宽,灵敏度降低,为了保证灵敏度和较短检[/font][font='黑体']测时间选择柱效高的3 μm粒径的氨基柱,在短时间内可以实现分离,检出限也达到要求。[/font][font='黑体']选择70%乙腈溶液作为流动相,样品中异构化乳糖和乳糖的分离度不好,无法检测;调整[/font][font='黑体']为乙腈为90% 时,异构化乳糖和乳糖可以分离,但出峰时间拖后至 30 min以后,峰型展[/font][font='黑体']宽,也不利于准确定量;选择80%乙腈可使样品中异构化乳糖和乳糖分离度达到要求,而且[/font][font='黑体']在20 min内出峰,峰型较好,可准确定量检测[/font][font='黑体']。[/font][font='黑体'] [/font][font='黑体'] [/font][font='黑体']酶法测定乳饮料中异构化乳糖的含量即[/font][font='黑体']乳饮料样品经过亚铁氰化钾和硫酸锌沉淀蛋白 质、β-D-半乳糖苷酶水解乳糖和异构化乳糖、葡萄糖氧化酶氧化葡萄糖等处理过程,用紫外分光光度计检测乳饮料中异构化乳糖的含量。该方法具有良好的回收率,为83.5%~92.7%,以及良好的重复性,相对标准偏差(RSD)为 4.04%~6.05%(n=6)。该方法检测结果准确且稳定性好,适用于乳饮料中异构化乳糖的检测[/font][font='黑体'][[/font][font='黑体']18]。[/font][font='黑体']在酶法中,酶的种类及沉淀剂的种类都会对异构化乳糖回收率造成影响。[/font][align=center][img]https://ng1.17img.cn/bbsfiles/images/2021/07/202107262012374933_8502_1608728_3.jpeg[/img][/align][align=center][font='黑体']表4[/font][font='黑体']不同沉淀剂对异构化乳糖回收率的影响[/font][/align][align=center][img]https://ng1.17img.cn/bbsfiles/images/2021/07/202107262012375761_4184_1608728_3.jpeg[/img][/align][align=center][font='黑体']表5[/font][font='黑体']不同酶的种类对异构化乳糖回收率的影[/font][font='黑体']响[/font][/align][font='等线'][size=13px]4. [/size][/font][font='等线'][size=13px]异构化乳糖的限量[/size][/font][font='等线'][size=13px][color=#333333]常见【异构化乳糖液】限量信息如下表6所示:[/color][/size][/font][table][tr][td][align=left][font='黑体'][color=#333333]类号[/color][/font][/align][/td][td][font='黑体'][color=#333333]食品名称[/color][/font][/td][td][font='黑体'][color=#333333]最大使用量[/color][/font][/td][td][font='黑体'][color=#333333]备注[/color][/font][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]01.03[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail9.html][font='黑体'][size=13px][color=#2d96cf]乳粉(包括加糖乳粉)和奶油粉及其调制产品[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]15.0 (g/kg)[/color][/size][/font][/td][td][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]01.03.02[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail11.html][font='黑体'][size=13px][color=#2d96cf]调制乳粉和调制奶油粉[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]15.0 (g/kg)[/color][/size][/font][/td][td][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]07.03[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail181.html][font='黑体'][size=13px][color=#2d96cf]饼干[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]2.0 (g/kg)[/color][/size][/font][/td][td][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]07.03.01[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail182.html][font='黑体'][size=13px][color=#2d96cf]夹心及装饰类饼干[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]2.0 (g/kg)[/color][/size][/font][/td][td][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]07.03.02[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail183.html][font='黑体'][size=13px][color=#2d96cf]威化饼干[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]2.0 (g/kg)[/color][/size][/font][/td][td][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]07.03.03[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail184.html][font='黑体'][size=13px][color=#2d96cf]蛋卷[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]2.0 (g/kg)[/color][/size][/font][/td][td][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]07.03.04[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail185.html][font='黑体'][size=13px][color=#2d96cf]其他饼干[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]2.0 (g/kg)[/color][/size][/font][/td][td][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]13.01[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail296.html][font='黑体'][size=13px][color=#2d96cf]婴幼儿配方食品[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]15.0 (g/kg)[/color][/size][/font][/td][td][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]13.01.01[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail297.html][font='黑体'][size=13px][color=#2d96cf]婴儿配方食品[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]15.0 (g/kg)[/color][/size][/font][/td][td][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]13.01.02[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail298.html][font='黑体'][size=13px][color=#2d96cf]较大婴儿和幼儿配方食品[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]15.0 (g/kg)[/color][/size][/font][/td][td][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]13.01.03[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail299.html][font='黑体'][size=13px][color=#2d96cf]特殊医学用途婴儿配方食品[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]15.0 (g/kg)[/color][/size][/font][/td][td][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]14[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail306.html][font='黑体'][size=13px][color=#2d96cf]饮料类[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]1.5 (g/kg)[/color][/size][/font][/td][td][font='黑体'][size=13px][color=#333333]固体饮料按稀释倍数增加使用量(14.01包装饮用水除外)[/color][/size][/font][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]14.02[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail311.html][font='黑体'][size=13px][color=#2d96cf]果蔬汁类及其饮料[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]1.5 (g/kg)[/color][/size][/font][/td][td][font='黑体'][size=13px][color=#333333]固体饮料按稀释倍数增加使用量(14.01包装饮用水除外)[/color][/size][/font][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]14.02.03[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail314.html][font='黑体'][size=13px][color=#2d96cf]果蔬汁(浆)类饮料[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]1.5 (g/kg)[/color][/size][/font][/td][td][font='黑体'][size=13px][color=#333333]固体饮料按稀释倍数增加使用量(14.01包装饮用水除外)[/color][/size][/font][/td][/tr][tr][td][font='黑体'][size=13px][color=#333333]14.03[/color][/size][/font][/td][td][url=http://www.foodcta.com/tjj/spxlDetail315.html][font='黑体'][size=13px][color=#2d96cf]蛋白饮料[/color][/size][/font][/url][/td][td][font='黑体'][size=13px][color=#333333]1.5 (g/kg)[/color][/size][/font][/td][td][font='黑体'][size=13px][color=#333333]固体饮料按稀释倍数增加使用量(14.01包装饮用水除外)[/color][/size][/font][/td][/tr][/table][font='黑体'][size=16px] [/size][/font][font='黑体'][size=16px] [/size][/font][font='黑体'][size=16px]表6[/size][/font][font='黑体'][size=16px]5[/size][/font][font='黑体'][size=16px].[/size][/font][font='黑体'][size=16px]异构化乳糖液的国家标准[/size][/font][font='黑体']食品安全国家标准[/font][font='黑体']GB1886.176—2016[/font][font='黑体']异构化乳糖液在2[/font][font='黑体']016[/font][font='黑体']年8月3[/font][font='黑体']1[/font][font='黑体']号发布,2[/font][font='黑体']017[/font][font='黑体']年1月1号实施。[/font][font='黑体']本标准代替GB8816—1988《食品添加剂 异构化乳糖液》。本标准与[/font][font='黑体'] GB8816—1988相比,主要变化如下:修改了感官要求 修改了异构化乳糖含量、果糖含量、[/font][font='黑体']半乳糖含量、乳糖含量、砷(As)的指标要求 删除了比重、折光率、大肠菌群的指标要求 增[/font][font='黑体']加了依匹乳糖含量、pH、灼烧残渣、大肠埃希氏菌的指标要求和检验方法 增加了鉴别试验[/font][font='黑体']方法 修改了异构化乳糖含量、果糖含量、半乳糖含量、乳糖含量的检验方法 部分检验方法[/font][font='黑体']的引用标准调整为最新发布的版本[/font][font='黑体']。[/font][font='黑体']该[/font][font='黑体']标准适用于以乳糖为原料,在加热条件下经碱或不经碱异构化制得的食品添加剂异构[/font][font='黑体']化乳糖液, 或再经干燥制成的固体产品。[/font][font='黑体'] [/font][font='黑体'] 感官要求应符合表7的规定。[/font][align=center][img]https://ng1.17img.cn/bbsfiles/images/2021/07/202107262012377853_3322_1608728_3.png[/img][/align][align=center][font='黑体']表7[/font][/align][font='黑体']理化指标应符合表8的规定。[/font][img]https://ng1.17img.cn/bbsfiles/images/2021/07/202107262012378664_191_1608728_3.png[/img][align=center][font='黑体']表8[/font][/align][font='黑体']微生物指标应符合表9的规定。[/font][img]https://ng1.17img.cn/bbsfiles/images/2021/07/202107262012379738_1039_1608728_3.png[/img][align=center][font='黑体']表9[/font][/align][font='黑体']附录:[/font][font='黑体']A.3 异构化乳糖、果糖、半乳糖、依匹乳糖、乳糖含量的测定 [/font][font='黑体']A.3.1 试剂和材料 [/font][font='黑体']A.3.1.1 水:符合 GB/T6682规定的一级水。 [/font][font='黑体']A.3.1.2 乙腈:色谱纯。[/font][font='黑体']A.3.1.3 异构化乳糖标准品、果糖标准品、半乳糖标准品、依匹乳糖标准品、乳糖标准品:纯度≥99.0%。[/font][font='黑体']A.3.1.4 磷酸缓冲液:称取无水磷酸二氢钠1.15g,用水溶解后定容至1000mL。[/font][font='黑体']A.3.2 仪器和设备 [/font][font='黑体']A.3.2.1 高效液相色谱仪,配备示差折光检测器、恒柱温及真空脱气系统。 [/font][font='黑体']A.3.2.2 色谱柱:以全多孔硅胶为填充剂的氨基柱(Φ4.6mm×15cm,填料粒径3μm)或其他等效的色谱柱。 [/font][font='黑体']A.3.3 参考色谱条件 A.3.3.1 流动相:乙腈∶磷酸缓冲液=82∶18。 [/font][font='黑体']A.3.3.2 流速:1.3mL/min。 A.3.3.3 柱温:40 ℃±1 ℃。 [/font][font='黑体']A.3.3.4 进样量:20μL。[/font][font='等线'][size=13px]5. [/size][/font][font='等线'][size=13px]总结[/size][/font][font='等线'][size=13px]综上所述,[/size][/font][font='等线'][size=13px]异构化乳糖是功能性低聚糖中少的双糖。其显著的益生特性和多样的生理功[/size][/font][font='黑体']能使异构化乳糖广泛应用于食品领域和医药领域[/font][font='黑体']。目前[/font][font='黑体']我国异构化乳糖的生产面临的问题是[/font][font='黑体']国内工业生产的异构化乳糖糖浆和晶体的纯度不高,生产成本较高等。它们极大地影响了异[/font][font='黑体']构化乳糖的广泛应用;再者我国将异构化乳糖应用于食品行业和饲料行业还处于起步阶段,[/font][font='黑体']异构化乳糖的使用还仅限于一些特殊产品中(如药品)。相信随着科学的进步发展,功能[/font][font='黑体']性[/font][font='黑体']异构化乳糖的生产技术会大幅提高且其[/font][font='黑体']在食品医疗等方面的[/font][font='黑体']应用也会受到我国研究者的广泛关注。[/font][font='黑体'][size=18px]参考文献:[/size][/font][font='黑体'][[/font][font='黑体']1] 许本发. 异构化乳糖促进肠道中双歧杆菌增值作用的观察[J]. 中国乳品工业, 1986, 4(14): 104-108.[/font][font='黑体'][2] 王凤仙, 刘博亚. 异构化乳糖的生理功能及在食品工业中的应用[J]. 中国食品添加剂, 2015(5): 179-182. DOI:10.3969/ j.issn.1006-2513.2015.05.022.[/font][font='黑体'][3] 黄萌萌, 王加启, 卜登攀. 牛奶乳果糖的研究进展[J]. 中国乳品工业, 2007, 35(6): 54-57. DOI:10.3969/j.issn.1001-2230.2007.06.014.[/font][font='黑体'][4] 杨 凯,薛江超,张天博,贾云虹,宋晓青[/font][font='黑体'],[/font][font='黑体']李朝旭[/font][font='黑体'].[/font][font='黑体']异构化乳糖在婴幼儿配方奶粉中的应用[J]. DOI:10.16172/j.cnki.114768.2017.10.021.[/font][font='黑体'][5][/font][font='黑体']王建中.异构化乳糖的性质及利用[/font][font='黑体']—[/font][font='黑体']双裂乳酸杆菌因子.[/font][font='黑体'][[/font][font='黑体']6]McSweeney P L H,Fox P F. Advanced dairy chemistry volume 3:lactose,water,salts and minor constituents[M]. New York:Springer Science+Business Media,LLC,2009.[/font][font='黑体'][7] Scientific Opinion on the substantiation of health claims related to lactulose and decreasing potentially pathogenic gastro intestinal microorganisms (ID 806)and reduction in intestinal transit time (ID 807)pursuant to Article 13(1)of Regulation (EC)No 1924/2006;EFSA Journal 2010;8(10):1806[/font][font='黑体'][8] othe M K,Maathuis A,Bellmann S,et al. Dose-dependent prebiotic effect of lactulose in a computer-controlled in vitro model of the human large intestine [J]. Nutrients, 2017,9( 7) : 767.[/font][font='黑体'][9] Tomoda T,Nakano Y,Kageyama T. Effect of yogurt and yogurt supplemented with Bifidobacterium and / or lactulose in healthy persons: a comparative study [J].Bifidobacteria and Microflora,1991,10( 2) : 123-130[/font][font='黑体'][10] Mizota T,Mori T,Yaeshima T,et al. Effects of low dosages of lactulose on the intestinal function of healthy adults [J].Milchwissenschaft,2002,57( 6) : 312-315.[/font][font='黑体'][11] Schumann,Christian. Medical,nutritional and technological properties of lactulose. An update[J]. European Journal of Nutrition,2002,41(1):17-25.[/font][font='黑体'][12][/font][font='黑体']张[/font][font='黑体']宗岩,侯万喜.异构化乳糖 - 新型婴儿食品添加剂 [J]. 中国乳品工业,1994,22(14):168-170.[/font][font='黑体'][[/font][font='黑体']13] Nagendra R,BASKARAN M V and Rao S V. Shelf life of spray dried infant formula supplemented with lactulose [J]. Journal of food processing and preservation,1995,19(4): 303-315[/font][font='黑体'] [14] 胡强, 徐红兵, 李水军, 等. 液相色谱-串联质谱法测定尿中乳果糖、 甘露醇和乳糖含量[J]. 中国现代医学杂志, 2008, 18(13): 1810-1813[/font][font='黑体'][15] 曾文芳, 时巧翠, 陈永欣, 等. [url=https://insevent.instrument.com.cn/t/3p][color=#3333ff]离子色谱[/color][/url]电化学测定牛奶中的乳糖和乳果糖[J]. 食品科学, 2006, 27(5): 205-207. DOI:10.3321/ j.issn:1002-6630.2006.05.043[/font][font='黑体'][16] 刘芳, 杨瑞金, 张文斌, 等. 薄层色谱法快速分析乳果糖[J]. 食品 与发酵工业, 2008, 34(1): 119-123. DOI:10.13995/j.cnki.11-1802/ ts.2008.01.029.[/font][font='黑体'][17] 任一平, 陈青俊, 黄百芬. 高效液相色谱示差折光法测定食品中的乳果糖[J]. 中国乳品工业, 1997, 25(4): 35-37[/font][font='黑体'][[/font][font='黑体']18] 刘丽君[/font][font='黑体'],[/font][font='黑体']李素琴[/font][font='黑体'],[/font][font='黑体']赵贞[/font][font='黑体'],[/font][font='黑体']岳虹[/font][font='黑体'],[/font][font='黑体']李翠枝[/font][font='黑体'],[/font][font='黑体']邵建波[/font][font='黑体'].[/font][font='黑体']高效液相色谱法测定乳粉中异构化乳糖的含量[/font][font='黑体'].[/font][font='黑体'] 1671-5187(2016)01-0011-03 DOI:10.15922/j.cnki.jdst. 2016.01.004.[/font][font='黑体'][[/font][font='黑体']19] 付云双[/font][font='黑体'],[/font][font='黑体']赵 贞[/font][font='黑体'],[/font][font='黑体']万 鹏*[/font][font='黑体'],[/font][font='黑体']温国艳[/font][font='黑体'],[/font][font='黑体']黄文强[/font][font='黑体'],[/font][font='黑体']徐 红[/font][font='黑体'],[/font][font='黑体']李翠枝[/font][font='黑体'],[/font][font='黑体']吕志[/font][font='黑体'].[/font][font='黑体'] 酶法测定乳饮料中异构化乳糖的含量. 1007-7871(2020)04-0035-04.[/font]
请问如何用高效液相色谱法(采用AGP手性色谱柱)检查酒石酸托特罗定左旋异构体?流动相中有少量磷酸盐,能否只用15%的正丙醇水溶液冲洗该手性色谱柱(Chiral-AGP)?谢谢!
1.1使用范围:鲜乳最大使用量(g/kg): 1.51.2使用范围:乳粉最大使用量(g/kg): 15.0简介 异构化乳糖,外观为白色结晶,甜度是蔗糖的0.6倍,是黏度低、热值低、易溶于水、性能稳定、安全性高、使用方便的一种新型低聚糖,也是一种有特殊保健功能的还原性二糖。工业上可以用干酪生产的副产物乳清中的乳糖在氢氧化钠作催化剂条件下进行加热使异构化,经离子交换、脱色、浓缩结晶等工序制取。在反应物中加入硼酸盐有助于异构化反应。 异构化乳糖对人体健康功能显著。人体肠道内有益菌为双歧杆菌,异构化乳糖是双歧杆菌生长最好的糖源,它在小肠内不被分解,移到大肠内可被所有双歧杆菌利用,使双歧杆菌增长占优势、抑制腐败细菌及病原菌的生长,对改变肠内菌丛、保持肠道正常功能、防病治病抗老化等起重要作用。 异构化乳糖防治便秘和防癌功效也很显著。肠内双歧杆菌增殖,pH值降低,促进了肠的蠕动,使粪便变软,排便数量及次数均可增加,并能使肠内易造成腐败菌生长的无用物在肠内滞留时间缩短,起到防止肠癌的作用。 用母乳喂养的婴儿比人工喂养的婴儿的肠道双歧杆菌占绝对优势(占总菌的90%以上),前者比后者抵抗力强,患病率低,生长发育好,因为母乳中有促进婴儿肠内双歧杆菌生长的因子。如在奶粉中加入0.5%的异构化乳糖,人工喂养婴儿,可使婴儿肠内双歧杆菌比例增加,接近母乳育儿,抵抗力增强,促进生长发育,降低患病率。 异构化乳糖可广泛用于保健食品中,如添加到乳饮料、碳酸饮料、果汁饮料、糖果、奶粉等食品中作膳食疗效食品,也可单独作医疗用品。 国家标准 食品添加剂 异构化乳糖液 GB 8816-88 Food additive Lactulose liquid 本标准适用于以乳糖为原料,以氢氧化钠为异构剂制得的异构化乳糖液。本品是双叉杆菌(bifidus)的增殖因子,主要用于鲜奶、奶粉、饼干等食品中,具有帮助消化吸收蛋白质、乳糖,产生维生素B组等功能。异构化乳糖中含有四种糖,即:乳酮糖、乳糖、 乳酮糖结构式半乳糖、果糖。其中起增殖双叉杆菌作用的是乳酮糖、又叫乳果糖、乳士糖、半乳糖基果糖甙。乳酮糖的结构式: 分子式C12H22O11 分子量342.30(按1983年国际原子量表) 技术要求 1 外观:一级品为黄色透明液体。二级品为棕色透明液体。 2 异 异构化乳糖液应符合的要求构化乳糖液应符合下列要求: 验收规则 1 异构化乳糖液应由生产厂质量检验部门进行检验,生产厂应保证所有出厂的异构乳糖液均符合本标准,每批出厂产品都应附有一定格式的合格证。 2 使用单位有权按照本标准规定的验收规则和试验方法检验所收到的异构化乳糖液是否符合本标准。 3 用清洁、干燥的玻璃管从每批10%的包装物中取出试样,取样时将玻璃管垂直插入容器内部,搅拌均匀,而后取样,每批取样总量不得少于200mL,将试样分别放入两只干净的磨口塞玻璃瓶中,混匀,粘贴标签。注明生产厂名称,批号,取样日期,一瓶送化验室分析,一瓶密封放在暗处保存2个月备查。 4 若检验结果中有一项指标不符合标准,应重新自两倍数量的包装件中取出异构化乳糖液进行复验。重新检验的结果中,即使只有一项指标不符合国家标准,应视为此批异构化乳糖液不能验收。 5 若供需双方对异构糖液质量发生异议,需要仲裁时,仲裁机构可由双方协商选定,仲裁应按照本标准规定的验收规则和检验方法进行。 包装、标志、贮存、运输 1 食品添加剂异构化乳糖液应装入食品用无毒聚乙烯塑料桶或玻璃瓶中,并外部用纸箱包装,每桶净重5kg,每箱净重20kg。 2 外包装应附有下列标志:生产厂名、产品名称、生产日期,质量标准、净重、批号、本标准编号及“食品添加剂”字样。 3 异构化乳糖液系食品添加剂,严禁与酸、碱、有毒物品及其他易腐蚀物品放在一起、在贮存与运输过程中应避免有毒物质污染,应存放在阴凉暗处,不准倒放。 4 二级品保存期:不得超过4个月。




 400-860-5168转0230
400-860-5168转0230
 留言咨询
留言咨询
 400-629-8889
留言咨询
400-629-8889
留言咨询
 400-860-5168转2472
留言咨询
400-860-5168转2472
留言咨询










 我要推广仪器
我要推广仪器
 下载APP
下载APP


