
 关注
关注
已关注
![]() 已认证
已认证
粉丝量 0
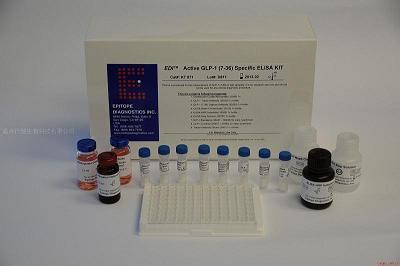
产品规格:100ug/200ug
产品用途:仅用于科研实验
本公司同类产品:
抗体的异质性。抗体的组成极为复杂,是由成千上万、多种多样的免疫球蛋白(Ig)分子所组成。这些Ig分子在形状、大小、结构以及氨基酸的组成和排列上,既相似,又有差别。由于有差别,它们的电泳活性就有很大的变化。
因为抗体具有与抗原决定簇相对应的结合部位(抗原结合簇),所以抗体与抗原的结合具有特异性。另一方面,抗体本身是一种蛋白质,具有本身的氨基酸组成、排列和立体结构,对异种动物来说,它又是抗原。各类Ig都具有可用血清学方法检出的抗原特异性,它们表现出不同的血清学类型。
其他产品:
①同种型特异性:同一种属所有个体共同具有的抗原特异性。人的Ig可分为5大类(IgM、IgG、IgA、IgD、IgE)、两个型(λ型和κ型)、以及若干亚类、亚型、群和亚群等。但是,抗体和抗原结合的特异性与抗体的类、亚类、型别等无关;
②同种异型特异性:同一种属(如人)的某些个体共有的抗原决定簇。这是由遗传基因决定的,某些重链和轻链(κ)的恒定区内,有个别氨基酸发生置换而形成Gm、Am、Km各型;
③独特型特异性:由单一B细胞克隆产生的Ig分子独有的抗原性。此决定簇在重链和轻链的可变区,特别是在可变区的超变区中。由于每一个体抗体形成细胞是由多克隆组成,所以独特型特异性为数极多。
任何一个正常人血清中的抗体都是含有成千上万的、多种多样的、具有各种独特型抗原性的免疫球蛋白分子混合物。κ链和 λ链可以和各类、各亚类配合。重链、轻链本身又有各种异型,每一克隆产生的Ig又具有自身的独特型。
抗体的多样性受 B细胞系统的遗传基因控制。肽链是由两个不同基因分别编码可变区或恒定区,恒定区的基因(C基因)是有限的,它虽然可以决定Ig分子的类别和亚类,为造成Ig分子多样性的原因之一,但是,造成免疫球蛋白分子多样性的主要原因却在于可变区的异质性,可变区是由V基因编码的,而V基因的数目仍不清楚。
对抗体多样性的遗传控制曾提出3种学说:
①种系学说(又称胚系学说):抗体形成细胞具有编码Ig分子的全部基因(即有限数量的C基因和未知数量的V基因,它是通过长期进化形成并通过生殖细胞从亲代传给子代。又称胚系学说;
②体细胞突变学说:在生殖细胞内只继承了数量有限的V基因,Ig分子多样性的形成,是由于体细胞在发育过程中发生突变或基因重组,从而产生许多不同的 V基因。体细胞突变可能对Ig分子的多样性发生重要作用;
③V区基因相互作用学说:Ig分子可变区是由V基因片段、J基因片段和D基因片段组成,V、D、J基因相互连接对Ig分子多样性的产生,是极为重要的。Ig的多样性不可能简单地归因于上述的某一学说,它可能与上述多种机制有关,也可能还与基因片段连接点上的连接多样性以及VL和VH链的不同配对有关。
更多![]()




企业名称
上海谷研实业有限公司
企业信息已认证
企业类型
信用代码
310112001219344
成立日期
2011-01-12
注册资本
100
经营范围
从事生物科技、医药科技领域内的技术咨询、技术服务、技术开发、技术转让,商务咨询(除经纪),电子商务(不得从事增值电信、金融业务),投资管理,建设工程(凭许可资质经营),化工原料及产品(除危险化学品、监控化学品、烟花爆竹、民用爆炸物品、易制毒化学品)、实验室设备、测量器材、酒店设备、环保设备、钟表、服装鞋帽、皮具箱包、一类医疗器械、电子产品、陶瓷制品的销售。【依法须经批准的项目,经相关部门批准后方可开展经营活动】
上海谷研实业有限公司
公司地址
上海松江区莘砖公路1290号
客服电话
公司名称: 上海谷研实业有限公司
公司地址: 上海松江区莘砖公路1290号 联系人: 黄经理 邮编: 201603
仪器信息网APP

展位手机站